Una tarea difícil III - Métodos de emparejamiento
Publicado el
por Gabriela.
AUTOR: Miguel Ángel Martín Espada
UNA TAREA DIFÍCIL III - MÉTODOS DE EMPAREJAMIENTO
1. Introducción.
Una vez seleccionados los reproductores, y a veces incluso antes, llega el momento de plantearse los emparejamientos que vamos a realizar para tratar de alcanzar esos objetivos que desde un principio nos hemos trazado y que, como ya sabemos, pretenden la obtención de ejemplares que nos deleiten con sus bellos y melódicos cantos discontinuos.
Al igual que vimos al tratar la selección de reproductores, hemos de diferenciar entre criadores noveles y criadores veteranos.
El margen de acción de los criadores noveles es más bien escaso, en sus inicios carecen de una base de conocimientos suficiente como para poder realizar la planificación de las parejas a reproducir, en la mayor parte de los casos simplemente seguirán el criterio de los criadores que les han proporcionado los ejemplares reproductores y elaborarán las parejas como aquellos les hayan aconsejado.
En los primeros años, aunque no introduzcamos ejemplares de otros criaderos y vayamos trabajando con la descendencia de nuestras primeras parejas, siempre en función de los resultados que vayamos obteniendo, y en tanto en cuanto adquiramos el bagaje de conocimientos mínimos necesarios, no podremos apartarnos del consejo de los veteranos, por ello seguiremos valiéndonos del asesoramiento de terceros para realizar los emparejamientos.
A medida que vayan pasando los años, que serán más o menos dependiendo exclusivamente de nuestra afición, interés y capacidad, iremos aprendiendo las reglas básicas e iremos ganando en independencia hasta que seamos autosuficientes y podamos planificar nosotros mismos nuestra política de emparejamientos; aunque conviene decir que no está de más confrontar nuestras ideas con las de otros aficionados cualificados, como se suele decir cuatro ojos ven más que dos.
En el presente escrito vamos a hacer una aproximación al tema de la elaboración de parejas desde el punto de vista tradicional de los manuales de canaricultura, selección, mejora y genética animal. No obstante, el criador de canarios de canto sabe que en este su particular arte debe tomar estas enseñanzas con gran cautela, ya que si bien en las variedades de color y postura es mucho lo que se ha avanzado, en canto seguimos en pañales. Y si lo pensamos bien no puede ser de otra forma, ya que si importantes son en todas las variedades los factores ambientales, en canaricultura de canto lo son más todavía, pues son tantos los elementos que podemos incluir en este apartado de la ecuación que es imposible controlarlos a todos; debemos conformarnos con poder actuar sobre unos pocos y, con todo, en muchas ocasiones estos factores decantan de tal modo la balanza, que el resultado final que obtenemos, puesto de manifiesto en el canto de nuestros canarios, no es siquiera remotamente indicativo de las cualidades genéticas que portan nuestros ejemplares.
En el llamado "Triangulo de la vida" de Walter, citado por autores tan importantes como Giorgio de Baseggio1, se da un paso más en el desarrollo de la tradicional ecuación fenotipo = genotipo + medio ambiente.
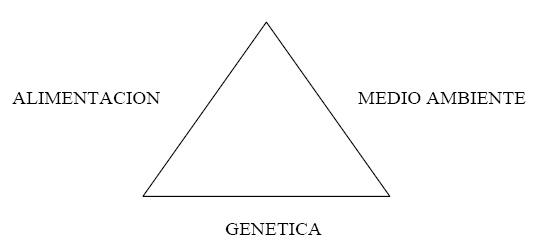
Como se puede observar, Walter2 da una crucial importancia a la alimentación, concediéndole un lugar propio, nosotros, hasta la fecha, hemos considerado la alimentación como un factor ambiental. La verdad es que la experiencia da la razón a Walter y la alimentación bien merece ese lugar de privilegio, siendo equiparada en importancia, por sí misma, al resto de factores ambientales y al genotipo. Walter defiende que para conseguir nuestros objetivos en la selección animal hemos de conseguir el equilibrio entre los tres factores que conforman los lados de su triángulo de la vida.
Como he apuntado anteriormente, los canaricultores de canto sabemos muy bien que esto es cierto, mediante un adecuado sistema de emparejamientos podemos acumular en nuestros ejemplares la información genética deseada, pero sin controlar los factores ambientales y la alimentación no se consigue nada.
En este artículo veremos cómo jugar con los emparejamientos para tratar de acumular en nuestros ejemplares las características genéticas deseadas, pero no perdamos de vista el resto de componentes de la ecuación3.

Cariotipo del canario en el que se pueden observar sus 80 cromosomas. Fuente: "O paporrubio"
2.- Parte General: Métodos de reproducción.
Una posible clasificación de los emparejamientos que podemos realizar es aquella que distingue entre consanguíneos y abiertos4.
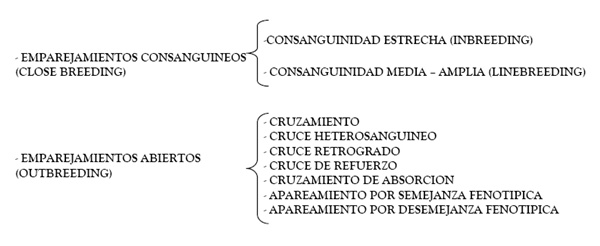
2.1 Emparejamientos Consanguíneos.
Como su nombre indica este método consiste en el emparejamiento de ejemplares que tienen uno o varios ascendientes comunes y que, por tanto, comparten, en mayor o menor medida, información genética procedente de ese tronco familiar común.
Para la reproducción en consanguinidad se viene utilizando tradicionalmente la denominada "Tabla de Felch". Dicha tabla representa las diferentes posibilidades de emparejamiento que tenemos partiendo de una pareja original, en principio no emparentada, y los resultados teóricos que se obtienen, desde el punto de vista genético, haciendo un emparejamiento u otro. Este sistema se basa en la genética mendeliana y posteriores avances de la ciencia genética han puesto de manifiesto fenómenos que constituyen excepciones a las leyes de Mendel y que alteran los resultados porcentuales de la Tabla, tales como puedan ser el crossing-over y el linkage, entre otros. Sea como fuere, hoy en día siguen siendo muchos los criadores que utilizan la Tabla de Felch como referencia orientativa a la hora de planificar los emparejamientos. En algunos textos la Tabla se representa mediante el esquema de emparejamiento de un macho con dos hembras e incluso con la posibilidad de un cruce de refresco.
A continuación reproduzco dos de las representaciones más habituales de la Tabla de Felch, este tipo de esquemas no suele precisar más comentarios dada su sencillez y evidencia.
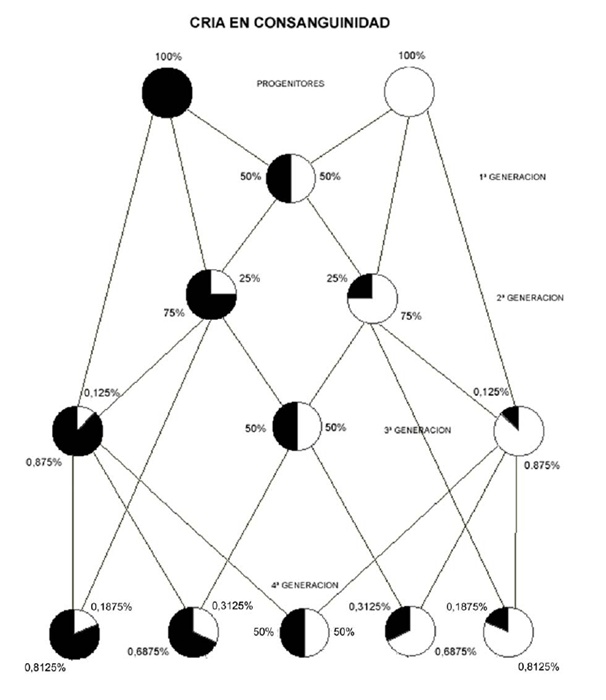
ESQUEMA DE EMPAREJAMIENTOS BASADO EN LA TABLA DE FELCH5
2.1.1 Emparejamientos en consaguinidad estrecha (Inbreeding).
Consiste en emparejar ejemplares emparentados en primer o segundo grado. Recordemos que cabe distinguir parentesco en línea recta (el existente entre individuos que descienden unos de otros: bisabuelos, abuelos, padres, hijos...) y parentesco colateral (el de individuos que no descienden unos de otros pero que proceden de un tronco común: hermanos, hermanastros, tíos, primos...). Para medir el grado de parentesco en línea recta entre dos ejemplares, se cuenta, excluyendo la del ejemplar del que partimos, el número de generaciones que hay entre ambos; y en caso de parentesco colateral, se cuenta el número de generaciones desde el primer ejemplar (excluyendo como en el caso anterior al ejemplar de partida) hasta el ascendiente común más próximo y después se sigue contando desde este último hasta llegar al segundo ejemplar, el número resultante será el grado de parentesco. Veamos un ejemplo práctico:

Siendo Z el ejemplar objeto de estudio, E y F, sus padres, tendrán un grado de parentesco lineal de 1º grado, mientras que A, B, C y D, los abuelos, lo tendrán de 2º grado.
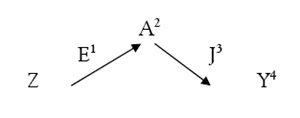
Veamos ahora otro árbol genealógico:
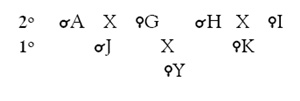
¿Existe parentesco entre Z e Y y, en caso de que lo haya, en qué grado? Bien, a la vista de los pedigrís resulta que ambos tienen un ascendiente común, el macho A, abuelo común (son, por tanto, primos hermanos, ya que sus respectivos padres eran hermanastros por parte de padre). Siguiendo las reglas expuestas vemos que de Z a A hay dos grados y de A a Y otros dos, por tanto entre Z e Y existe un parentesco colateral de cuarto grado.
Atendiendo a las explicaciones de los párrafos precedentes la consanguinidad estrecha es la practicada mediante el emparejamiento entre padres-hijos (primer grado), nietos-abuelos (segundo grado) o hermanastros (2º grado).
Resulta evidente que se trata de los emparejamientos de consanguinidad más elevada, o, dicho de otra manera, aquellos en los que los reproductores tienen en común un mayor número de genes y, por tanto, mayor posibilidad de conseguir homocigosis para los caracteres deseados.
Yo suelo englobar a este tipo de consanguinidad dentro de lo que denomino consanguinidad directa, ya que suele hacerse para fijar las características de ejemplares concretos, mediante el emparejamiento de aquellos con sus hijos y nietos.
Aconsejo a los criadores noveles que no practiquen este tipo de consanguinidad durante más de dos generaciones seguidas, el paso de ese límite debe hacerse tan solo por criadores experimentados. La consanguinidad estrecha es la forma más rápida de fijar los caracteres deseados pero también de que afloren los no deseados de carácter recesivo, como por ejemplo los letales y subletales. En caso de que los ejemplares con los que iniciamos nuestro trabajo sean portadores de información genética negativa, la consanguinidad estrecha puede hacerla aflorar ya en los primeros emparejamientos, lo que probablemente nos haga tener que abandonar a las primeras de cambio. Veremos más adelante una forma de evitar que pase esto.
2.1.2 Emparejamientos en consaguinidad media-amplia (Linebreeding).
Una forma de aprovechar las ventajas de la cría en consanguinidad, y a la vez tratar de retrasar las consecuencias negativas que, tarde o temprano, ésta implica, es el emparejamiento de ejemplares en consanguinidad media-amplia, o como se dice en inglés Linebreeding.
Consanguinidad media es el parentesco entre 3er y 4º grado y amplia el parentesco entre 5º y 10º grado. A partir de 11º grado se considera que el parentesco es ya tan lejano que no cabe hablar ya de emparejamiento consanguíneo.
Lógicamente, cuanto más lejano sea el grado de parentesco menores serán las posibilidades de que se encuentren los genes comunes o de que se produzca la combinación genética deseada y también menores las posibilidades de cumplir el objetivo deseado. Entre los expertos suele decirse que la consanguinidad amplia es poco efectiva en la cría selectiva de alto nivel.
En la práctica la consanguinidad estrecha se utiliza para fijar genéticamente las características deseadas y la amplia-media para mantenerlas o conservarlas, más o menos puras, durante el mayor tiempo posible sin introducir ejemplares ajenos.
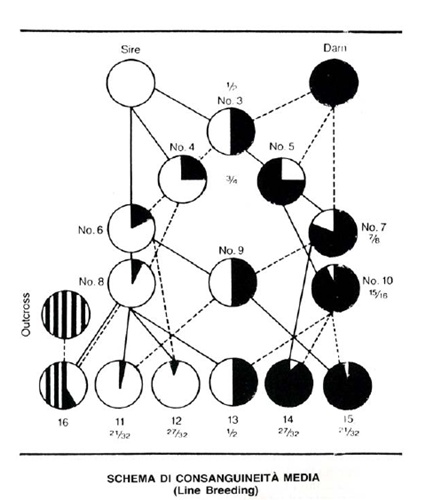
Esquema en el que se pueden ver emparejamientos en inbreeding, linebreeding y "refresco". Fuente: G. de Bassegio (obra citada en las anotaciones).
Dentro de la consanguinidad media-amplia y a efectos puramente didácticos, distingo entre emparejamientos en línea recta (bisabuelos-biznietos, tatarabuelos-tataranietos…), que englobo dentro de la consanguinidad directa vista en el punto anterior (búsqueda de las características genéticas de ejemplares concretos), y los emparejamientos en línea colateral, a los que denomino consanguinidad indirecta ya que suelen buscar las características genéticas compartidas por el tronco familiar en su conjunto, más que las características de ejemplares concretos.
2.2. Emparejamientos abiertos (outbreeding).
Este tipo de emparejamientos se realiza entre ejemplares no emparentados o de parentesco lejano o insignificante. Como veremos seguidamente se suele
realizar para refrescar una línea consanguínea, para corregir desviaciones mediante compensación (en caracteres de genética cuantitativa) o para incidir en determinadas características mediante cruces por semejanza. Lo importante de este tipo de cruces es que aumentan la variación genética y la heterocigosis.
Algunos de los emparejamientos que siguen se citan simplemente para dejar constancia de los mismos, ya que en nuestro caso o no son necesarios o, en ocasiones, la oportunidad de hacerlos corresponde a criadores con muchos años de experiencia y por motivos muy concretos.
2.2.1. Cruzamientos.
Utilizar el término cruzamientos en sentido estricto es, no cabe el menor género de dudas, peliagudo. Para muchos el término raza adquiere un significado cuasi sagrado, la pureza de la raza o de sangre son expresiones de constante uso en la cría deportiva de animales domésticos, pero los técnicos en la materia son más prácticos.
Dalton , por ejemplo, dice que "la mayoría de las razas puras de hoy fueron ayer cruzamientos". Suele creerse que una vez fijadas y homologadas las características genéticas, fenotípicas y, a veces, etológicas o conductuales, que determinan que una población animal pueda ser considerada como raza ésta se convierte en un compartimiento genético estanco en el que no cabe introducir nueva información genética por la vía de cruces con otras especies o razas de su misma especie. Pero de la lectura de los manuales de zootecnia, mejora y genética animal no se desprende esto ni mucho menos.
En los manuales se distingue claramente entre mestizaje y cruzamiento. Mestizaje es el fruto de un accidente o de apareamientos sin un programa o finalidad concreta; y el cruzamiento el fruto de un apareamiento programado. Los expertos admiten el cruzamiento tanto con la finalidad de crear nuevas razas como con la de introducir características concretas para su mejora en razas establecidas (no nos compete hablar aquí de los cruzamientos para obtener ejemplares cruzados de mayor valor comercial que las razas puras originarias por separado).
Evidentemente, los cruzamientos entre especies o razas diferentes son sumamente peligrosos, puesto que si no son realizados de forma selectiva y con el debido control pueden producir el efecto contrario al deseado: el bastardeo de la raza e incluso su desaparición. Desaconsejamos
completamente este tipo de cruces, a pesar de que vamos a ver ejemplos de los mismos a modo divulgativo.
Podemos distinguir los siguientes tipos de cruzamientos:
- Entre especies diferentes.
- Entre razas diferentes.
- Entre estirpes, líneas, o familias diferentes.
.
a) Entre especies diferentes:
Se realizan para introducir características concretas de una especie en otra, en nuestro caso en el canario (normalmente se trata de caracteres de genética cualitativa, es decir, controlados por una o pocas parejas de genes). El procedimiento a seguir una vez obtenido el híbrido fértil, el f1, es retrocruzarlo, seleccionando la característica deseada, durante un mínimo de tres generaciones con canarios de la raza parental, la cuarta generación (los r-3, que no f-4, son considerados ya canarios puros). Veámoslo en este gráfico:
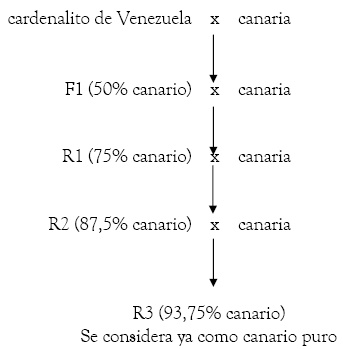
En Canaricultura de Color han dado lugar a las variedades de factor rojo y a la categoría mosaico (cruce entre Cardenalito de Venezuela -carduelis cucullata- y canario); a través de este tipo de cruzamientos con otros spinus sudamericanos (como el Negrito de Bolivia -carduelis atrata-) se está tratando de conseguir el canario negro y, recientemente, a través del cruzamiento con el carduelis magellanicus se ha conseguido el color "jaspe"6. Asimismo, se ha cruzado con verdecillo (serinus serinus) para mejorar el diseño melánico.
En Postura también se ha utilizado el cruce entre especies. Algunos sospechan, aunque no está demostrado, que el Lizard es fruto de cruce con otra especie. En las razas miniatura, como la Raza Española, se ha cruzado con verdecillo (serinus serinus) para minorar más el tamaño; tal ha sido el abuso y las nefastas consecuencias de este tipo de cruces en el Raza Española que hoy en día existe un grave problema entre criadores y jueces de esta raza, ya que en los concursos se miran con desconfianza los ejemplares de color verde, toda vez que los expertos dicen que son más los inconvenientes que las ventajas en este tipo de cruce.
Por último, en canto, Drove7 pensaba que en el origen del moderno canario Harzer, había intervenido la hibridación con verdecillo, verderón y verderón serrano, se trataba de una hipótesis y Drove afirmaba haber realizado cruces con estas especies, destacando el citado maestro los resultados obtenidos con el verdecillo. En el seno del Timbrado Español tengo constancia de que algunos criadores han intentado sin resultado el cruce con verderón serrano, con verdecillo, así como con cantor de África (serinus leugopygius) y otros serinus africanos como el canario del cabo (serinus canicollis) o el canario de vientre amarillo (serinus flaviventris), sin que, por lo que se comenta, el resultado haya sido satisfactorio.
En este sentido, yo mismo he de reconocer que en su día me planteé hacer un experimento con cantor de África, pero después de analizar el canto de este primo del canario y reflexionar sobre si podía aportar algo al canario discontinuo, llegué a la conclusión de que, hoy por hoy, dado el grado de desarrollo genético y la perfección de nuestros canarios discontinuos, este tipo de cruces no nos podía aportar nada que no pudiéramos conseguir dentro de nuestros propios canarios.
Otra cosa es utilizar este tipo de cruces entre especies para estudiar los mecanismos hereditarios del canto, pero con ánimo puramente científico y siendo el experimento el fin en sí mismo y no la introducción de características ajenas en nuestros canarios; hecho este, reitero, que, humildemente, pienso que no nos puede aportar nada hoy por hoy y sí un bastardeo de peligrosas e imprevisibles consecuencias.
b) Entre razas diferentes.
Este tipo de cruces se realiza en Canaricultura principalmente para introducir características de una raza en otra, o para crear nuevas razas. En las revistas especializadas8 podemos encontrar frecuentes referencias a este tipo de cruces, sobre todo en Postura; es sumamente educativo leer cómo siguiendo una compleja planificación de cruces entre dos o incluso tres razas y con una exhaustiva selección se han conseguido muchos de los canarios de Postura que actualmente compiten en nuestros concursos. Hasta tal punto han proliferado las razas de Postura, que la COM se ha visto obligada a tomar medidas para endurecer el procedimiento de homologación de nuevas razas, ya que se afirma que muchas razas aún guardan tal parecido entre sí que en los concursos pueden confundirse incluso por jueces expertos (es conocido y notorio que en algunas razas, según el fenotipo obtenido, los criadores presentan los ejemplares a concurso con una denominación u otra).
En canto es mucho lo que se ha especulado en el origen de las diferentes razas respecto al tema de los cruces interraciales, sin que pueda afirmarse fehacientemente nada al respecto.
Al igual que dije al hablar de los cruces entre especies, me parece correcto experimentar con este tipo de cruces para estudiar los mecanismos de la herencia en el canto, pero con esa única y exclusiva finalidad. La experiencia de los enjuiciamientos en los concursos demuestra que los cruces entre variedades de canto siempre terminan aflorando.
Cierto es que hay criadores habilidosos que han sido capaces de introducir las características deseadas siguiendo el esquema de los cruces por absorción, que veremos, y una rigurosísima selección; pero la cantidad de ejemplares que de forma poco inteligente y temeraria han ido vendiendo o regalando por el camino a criadores menos hábiles y la consanguinidad entre ejemplares "no depurados" han hecho aflorar las características de la raza ajena. En los últimos años ha aumentado peligrosamente en algunas zonas el número de ejemplares descalificados por canto Malinois, tanto entre los criadores de Timbrado clásico e intermedio como entre los criadores de canarios discontinuos.
c) Entre estirpes, líneas, o familias diferentes.
En genética de poblaciones una raza puede dividirse en9:
- Variedades: En función de diferencias morfológicas que no afectan a la conformación o formato básico que caracteriza y define a la raza como tal.
- Estirpe: "Población cerrada de animales de una raza o variedad, que ha sido creada por algún criador a base de reproducirla con individuos pertenecientes a ella misma, sin introducir material extraño".
- Línea: "La línea dentro de la estirpe está constituida por la reproducción, en cada generación, de un grupo de animales que se han ido eligiendo por alguna razón determinada, independientemente de la reproducción del resto de la estirpe".
- Familia: Diferentes ramas parentales dentro de una Línea.
Hay que reconocer que dentro del canario discontinuo, y de la canaricultura de canto en general, esta terminología no se usa de una forma demasiado acertada desde el punto de vista técnico, ya que se confunden los conceptos variedad, estirpe y línea. Resultaría polémico a la par que estéril entrar en este punto, digamos, simplemente y para salir lo más airadamente posible del brete, que dentro del canario discontinuo las diferentes subtendencias de canto10 podrían representar variedades; dentro de esas variedades, cuando un grupo de criadores desarrolla una familia trabajando en consanguinidad y sin introducir ejemplares ajenos hasta el punto de conseguir un número de ejemplares más o menos elevado con características fenotípicas afines podríamos hablar de estirpe, siendo la población concreta de cada criadero, con sus características específicas y diferenciadas, su línea y dentro de cada criadero cada rama de trabajo consanguínea diferenciada una familia. Sé que es quizás un tanto forzado, pero lo cierto es que hoy por hoy estos conceptos son de difícil aplicación a nuestro caso concreto. Toda vez que tampoco seguimos al pie de la letra las técnicas de reproducción utilizadas en mejora animal, al estar pensadas para poblaciones elevadas de animales y no para las reducidas cantidades que nosotros tenemos en nuestros criaderos.
Este tipo de cruces se realizaría entre variedades diferentes de la misma raza, entre estirpes diferentes dentro de la misma variedad, entre líneas diferentes dentro de la misma estirpe o entre familias dentro de una misma línea y sin que los ejemplares cruzados tuviesen entre sí un parentesco por debajo del 11º grado, ya que si no estaríamos hablando de consanguinidad amplia y no de un cruce abierto. Lógicamente estos conceptos están pensados para explotaciones ganaderas y resultan difíciles de aplicar a
nuestros criaderos. En nuestros aviarios la consanguinidad media entre los ejemplares difícilmente baja de 5º ó 6º grado.
2.2.2. Cruce heterosanguíneo.
Es el que llamamos coloquialmente cruce de refresco o de introducción en nuestro criadero de "sangre (genes) nueva".
2.2.3. Cruce retrogrado.
Cruce del f1 de un cruzamiento con uno de sus progenitores de raza pura.
2.2.4. Cruces de refuerzo y absorción.
Siguiendo a Dalton agrupamos estos dos tipos de cruces dada su semejanza.
"El cruce de refuerzo se realiza cuando un criador retrocede hasta la fuente genética original de la raza o estirpe en busca de nuevo material genético". Este tipo de cruce es el realizado por los criadores que cruzan con canario silvestre con la intención de aportar al canto de sus ejemplares matices de aquél y no para refrescar, que es la otra finalidad de este tipo de cruces.
"El cruzamiento de absorción es aquel en el que una raza se va transformando en otra mediante cruzamientos continuados".
Imaginemos que un criador adquiere una pareja canarios de una raza extranjera muy difícil de obtener y, por tanto, de elevado coste y quiere conseguir aumentar el número de ejemplares de la misma por encima de las posibilidades de reproducción de una sola pareja. Cruzaría el macho puro, al que vamos a llamar de raza A, con hembras de otra raza, B, según el siguiente esquema y con una rigurosa selección fenotípica buscando siempre las características de la raza A:

Con los cruces de refuerzo se sigue el mismo esquema de cruzamientos. El problema empieza cuando en segunda generación ya se obtienen buenos resultados. Los criadores se suelen olvidar de que el R1 es todavía un medio y no un fin en sí mismo; y que, por muy bueno que sea, hay que seguir cruzándolo con ejemplares de la raza parental que se quiere reforzar durante un mínimo de otras dos generaciones para llegar al R3, so pena de fijar ejemplares con un exceso de genes de la otra raza que trabajados en consanguinidad pueden dar lugar a una población bastardeada, en mayor o menor medida, cuyas características no se corresponderán con ninguna de las dos razas originales.
Esto es lo que ha pasado en el seno de nuestro canario con los cruces con canario silvestre. Los buenos resultados obtenidos por muchos criadores ya con los R1 (mal lamados F2), han hecho que los R1 y R2 se crucen entre sí y que incluso sean trabajados en consanguinidad estrecha, consiguiendo que en generaciones sucesivas el porcentaje de genes silvestres aumente y en lugar de avanzar hacia el canario doméstico lo que fijamos es una población mestiza en la que se pierden los avances canoros conseguidos durante generaciones de trabajo selectivo en el canario doméstico.
Para evitar esto hemos de seguir al pie de la letra los esquemas de cruzamiento previamente vistos.
En el cruce de refuerzo con el canario silvestre, tal y como ya hemos apuntado, podemos pretender dos cosas:
a) Aportar matices canoros.
b) Refrescar poblaciones consanguíneas.
Respecto a los cruces, con independencia de lo que pretendamos, podemos seguir dos posibilidades, como decía Rafael Martínez Bouzo, la rápida y la lenta. Estas dos vías se basan en la teoría macho da hembra y hembra da macho, seguida al pie de la letra por criadores como Santiago Iso Gracia. Como expliqué en "Consideraciones sobre la herencia en los canarios de Canto"11 yo voy más allá en mi hipótesis de trabajo y parto de la premisa de que los genes que rigen el patrón genético de canto son de genética cualitativa y de carácter ligado al sexo (es decir, que se ubican en el cromosoma sexual masculino, el Z). Además también trabajo con la hipótesis de que al hacer un emparejamiento hay tres factores principales, desde el punto de vista genético, que influyen en el canto y que se transmiten de forma independiente:
1) Aparato de Canto, el instrumento, que marcará la voz: controlado por muchos pares de genes (genética cuantitativa) autosómicos (no ligados al sexo).
2) Patrón innato de Canto, el que marca la predisposición hacia un tipo u otro de giros: de genética cualitativa y ligado al sexo.
3) Inteligencia, talento o habilidad del canario como intérprete: creo que los genes que lo determinan son de genética cuantitativa y herencia autosómica aunque todavía tengo mis dudas.
Nos centraremos en la presente exposición en el factor Patrón innato de Canto, ya que la experiencia me dice que es el más fácil de manejar y la que me hace pensar que es de genética cualitativa y de carácter ligado al sexo12:
- Vía rápida: Distinguimos entre porcentaje teórico general y herencia del patrón genético del canto.
Transmisión del patrón genético de canto contenido en el cromosoma Z:
1ª Generación: En rojo los cromosomas del canario silvestre, en azul los de la hembra doméstica, el cromosoma w, que determina el sexo femenino al igual que acontece en color no contendría información genética canora:
| CROMOSOMAS | Z | W |
| Z | Z Z | Z W |
| Z | Z Z | Z W |
Podemos observar que todos los machos serían heterocigóticos canoramente, su patrón genético de canto sería mitad doméstico, mitad silvestre; mientras que las hembras, al haber heredado el cromosoma Z del silvestre, serían portadoras sólo de la información genética canora de su progenitor.
El hecho de que todos los machos sean canoramente heterocigóticos explica las sabidas carencias de los F1.
2ª Generación: Partimos de la premisa de que no se ha producido ningún tipo de recombinación génica ni cromosómica que pudiera afectar a la información contenida en los cromosomas sexuales Z, provocando intercambio de información entre los dos cromosomas Z durante la fase de meiosis13.
| CROMOSOMAS | Z | W |
| Z | Z Z | Z W |
| Z | Z Z | Z W |
En la segunda generación, vemos que el retrocruce de un macho F1 con hembra doméstica (emparentada o no), complica las combinaciones teóricas de los R1.
Desde un punto de vista teórico vamos a tener las siguientes combinaciones canoras:
- Machos: Z Z, a pesar de tener un porcentaje teórico de 75% de genes domésticos y 25 % de genes silvestres, en lo canoro sigue siendo un ejemplar heterocigótico que en condiciones ambientales ideales seguiría cantando de forma parecida a los F1, si bien puede dar la impresión de emitir un canto más o menos próximo al doméstico en función de los otros dos factores principales (capacidad morfofisiológica -aparato de canto- y habilidad o inteligencia canora).
- Machos: Z Z, a pesar de tener un porcentaje teórico de 75% de genes domésticos y 25 % de genes silvestres, en lo canoro es un ejemplar homocigótico que en condiciones ambientales ideales emitiría un canto doméstico, si bien puede dar la impresión de emitir un canto más o menos asilvestrado en función de los otros dos factores principales (capacidad morfofisiológica -aparato de canto- y habilidad o inteligencia canora).
- Hembras: Z W, a pesar de tener un porcentaje teórico de 75% de genes domésticos y 25 % de genes silvestres, en lo canoro aportará el patrón canoro del silvestre.
- Hembras: Z W, a pesar de tener un porcentaje teórico de 75% de genes domésticos y 25 % de genes silvestres, en lo canoro aportará el patrón canoro del doméstico.
La complicación añadida en los machos R1 es que al poder tener en un mismo voladero ejemplares con combinaciones genéticas canoras diferentes no vamos a saber a ciencia cierta el genotipo canoro de nuestros ejemplares ya que fenotípicamente emitirán, normalmente, el canto de los ejemplares dominantes o compondrán una canción intermedia, fruto de la interacción canora entre ejemplares de combinación genética diferente. Por lo que podemos tener ejemplares de canto doméstico genéticamente heterocigóticos y ejemplares asilvestrados genéticamente domésticos. En el caso de las hembras tenemos un 50% de posibilidades.
Esto explica que los R1 puedan salirnos campeones o, por el contrario, casi tan asilvestrados como los F1. La única forma de saber si un macho R1 es homocigótico o heterocigótico es cruzarlo con sus propias hijas. Si salen todos los ejemplares con características canoras domésticas el R1 es homocigótico, si nos salen ejemplares de canto asilvestrado el R1 es heterocigótico. Esto no es teoría, es una conclusión fruto de la experimentación con ejemplares R1 y R2.
Lo dicho en los párrafos precedentes vale tanto para los R3 como para los R4, ya que podemos alejarnos genéticamente del silvestre pero seguir teniendo ejemplares de canto asilvestrado por la herencia de cromosomas Z de origen silvestre.
Incluso en el caso de contar con ejemplares con patrón genético doméstico, nos podemos encontrar con atavismos o rebotes de genes silvestres autosómicos que incidan en las capacidades morfológicas y fisiológicas y doten al canto de una patina de asilvestramiento fenotípico. Tal es la complejidad y dificultad de trabajar con ejemplares procedentes de cruces con canario silvestre y el motivo por el que sólo debería hacerse por criadores experimentados.
- Vía lenta:
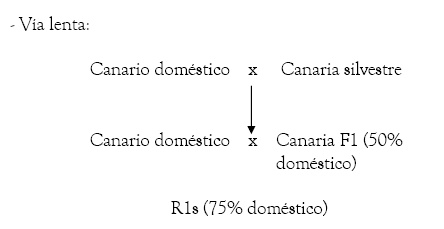
Desde de los R1, se pueden utilizar tanto los machos como las hembras, que deberán ser cruzados durante dos generaciones más con ejemplares domésticos (hasta llegar a los R3).
Apuntar que hasta hace no muchos años, ante la falta de canarias silvestres, de la imposibilidad de conseguir criar con las que se tenían, o simplemente por la falta de unas mínimas garantías canoras, se utilizaban principalmente las hembras F1 hijas de macho silvestre y hembra doméstica en lugar de canarias silvestres puras, motivo por el que se denominaba a este sistema vía lenta, ya que los resultados positivos no solían verse antes de los R2, mientras que con la vía rápida o con el uso de canaria silvestre, podían salir ya ejemplares R1 de calidad.
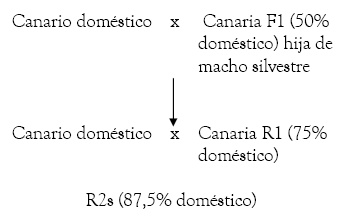
Desde de los R2s, se pueden utilizar tanto los machos como las hembras, que deberán ser cruzados durante una generación más con ejemplares domésticos (hasta llegar a los R3).
Transmisión del patrón genético de canto contenido en el cromosoma Z:
1ª Generación: En rojo los cromosomas del canario silvestre, en azul los del doméstico, el cromosoma w, que determina el sexo femenino al igual que acontece en color no contendría información genética canora:
| CROMOSOMAS | Z | W |
| Z | Z Z | Z W |
| Z | Z Z | Z W |
Podemos observar que todos los machos serían heterocigóticos canoramente, su patrón genético de canto sería mitad doméstico, mitad silvestre; mientras que las hembras, al haber heredado el cromosoma Z del doméstico, serían portadoras sólo de la información genética canora de su progenitor.
El hecho de que todos los machos sean canoramente heterocigóticos explica las sabidas carencias de los F1 o de los R1 hijos de hembra F1 hija de macho silvestre.
2ª Generación: Partimos de la premisa de que no se ha producido ningún tipo de recombinación génica ni cromosómica que pudiera afectar a la información contenida en los cromosomas sexuales Z, provocando intercambio de información entre los dos cromosomas Z durante la fase de meiosis.
| CROMOSOMAS | Z | W |
| Z | Z Z | Z W |
| Z | Z Z | Z W |
Con este sistema vemos que todos los ejemplares R1, o R2 hijos de hembra F1 de macho silvestre, son portadores del cromosoma Z doméstico; aunque como se ha indicado anteriormente nos podemos encontrar con atavismos o rebotes de genes silvestres autosómicos que incidan en las capacidades morfológicas y fisiológicas y doten al canto de una patina de asilvestramiento fenotípico, sobre todo en los R1, R2 y R3.
Conclusión: Si lo que queremos es aportar matices asilvestrados trataremos de trabajar con ejemplares heterocigóticos (ZZ) buscando que se produzca una recombinación cromosómica que de lugar a un cromosoma Z con información doméstica y silvestre, si bien con predominio de la primera, ya que como decía también Rafael Martínez Bouzo "el silvestrismo debe introducirse en nuestro canario con cuentagotas, de lo contrario deja de ser canto español" (cita textual). Ejemplos suficientes hay en nuestros canarios de que esto se produce.
Por el contrario, si sólo queremos refrescar trabajaremos siempre con ejemplares homocigóticos domésticos (ZZ).
2.2.5. Apareamientos por semejanza fenotípica.
Consisten en emparejar ejemplares por su semejanza fenotípica respecto a las características que queremos potenciar.
2.2.6. Apareamientos por desemejanza o compensación.
Consisten en el apareamiento de dos ejemplares de características diferentes con el fin de que éstas se compensen y eliminar o atenuar así las características no queridas.
3. Parte especial. Conclusiones.
En el punto anterior hemos hecho una aproximación genérica a los diferentes sistemas de emparejamiento, pero al criador lo que le interesa es saber concretamente cómo tiene que proceder en su caso concreto.
Es evidente que el tipo de emparejamientos a realizar va depender de muchos factores. Un principiante querrá saber principalmente cómo iniciar y fijar una línea, más tarde querrá saber cómo mantenerla, cómo introducir nuevas características y, en su caso, como refrescarla. A medida que nos hacemos como criadores nuestras inquietudes van variando y ganando en complejidad. Es, pues, difícil contentar a todos los criadores a la vez.
Con todo, vamos a intentarlo partiendo de una serie de pautas básicas. Tengamos presente antes de hacer los emparejamientos:
1º) Tener claro nuestro objetivo: Por qué hacemos esos emparejamientos y no otros y para qué.
2º) Al hacer un emparejamiento hay tres factores principales, desde el punto de vista genético, que influyen en el canto y que se transmiten de forma independiente:
a) Aparato de Canto, el instrumento, que marcará la voz: controlado por muchos pares de genes (genética cuantitativa) autosómicos (no ligados al sexo).
b) Patrón innato de Canto, el que marca la predisposición hacia un tipo u otro de giros: de genética cualitativa y ligado al sexo.
c) Inteligencia, talento o habilidad del canario como intérprete: ¿genética cuantitativa y herencia autosómica?
3º) Seleccionar los reproductores y realizar los emparejamientos procurando tener cierta polivalencia para que en caso de que no podamos realizar, por el motivo que fuere (muerte de uno de los reproductores, esterilidad, peleas, etc.), alguno de los emparejamientos existan otros ejemplares en el criadero con los que podamos utilizar los componentes de la pareja fallida con las mismas garantías de conseguir el fin perseguido.
Dicho de otra manera, tener un plan B; todos los años tenemos que replantearnos algún emparejamiento por diferentes motivos, se trata de tener este hecho presente desde el principio y haber previsto las posibles alternativas para evitar la improvisación, de tan nefastas consecuencias cuando de realizar un trabajo selectivo serio se trata.
4º) Seguir como referencia orientativa un esquema de emparejamientos, por ejemplo:
Esquema tradicional simplificado de cría en consaguinidad:
1ª Generación: Emparejamiento de un macho (1) con dos hembras (2 y 3), en principio no consanguíneos.
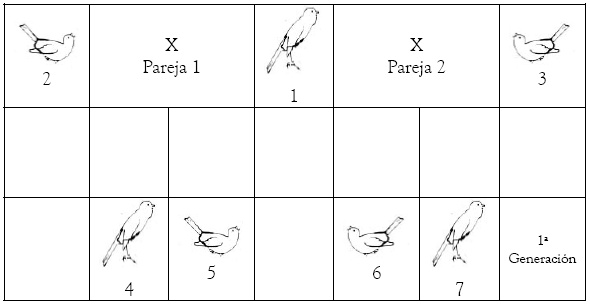
2ª Generación: Tenemos las siguientes opciones: Emparejamiento del macho 1 con dos de sus hijas (5 y 6), del macho 4 con su madre (2) y con una hermanastra (6) y del macho 7 con su madre (3) y con una hermanastra (5). Incluso podemos hacer emparejamientos no consanguíneos (macho 4 con hembra 3 y macho 7 con hembra 2).
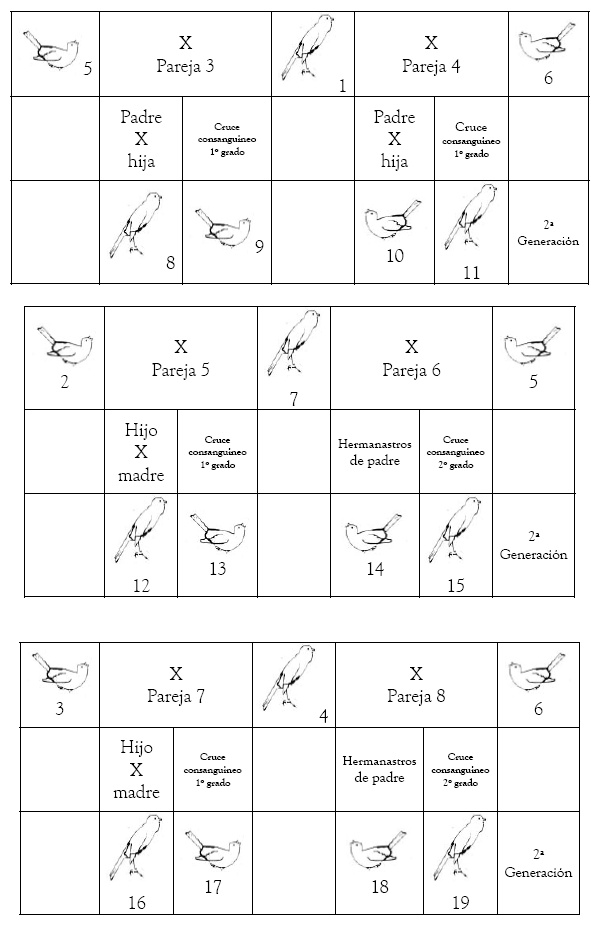
3ª Generación: las posibilidades de emparejamiento se multiplican de forma considerable. Podemos hacer cruces de abuelos/las con nietos/as (2º grado), padres/madres con hijos/as (1º grado), hermanastros (2º grado), tíos/as con sobrinos/as (3º grado) y entre primos (4º grado). También podemos hacer cruces no consanguíneos entre la descendencia de la hembra 2 no emparejada con la descendencia de la hembra 3 y esta última y viceversa.
Partiendo de un macho y dos hembras y en tan solo tres años, si los resultados han acompañado y los objetivos que nos hemos marcado se han ido cumpliendo tenemos un margen de maniobra considerable que nos permite hacer emparejamientos de mantenimiento de lo conseguido sin abusar en demasía de la consanguinidad (consanguinidad media y amplia) y sin tener que meter pluma ajena a nuestro criadero.
Lo normal es que en unos años consigamos una gran homogeneidad genética entre nuestros ejemplares, a medida que vayamos ganando en homocigosis iremos perdiendo margen de mejora hasta llegar a lo que los expertos llaman plateau o meseta consanguínea, para posteriormente iniciar la depresión consanguínea provocada por la pérdida de variedad genética y la acumulación de información genética negativa (genes letales y subletales, genes responsables de pérdida de vitalidad y capacidad funcional, etc.). Yo soy partidario de mantener cierto grado de variedad genética en nuestro criadero, a través del control de los emparejamientos. Recomiendo tener un tronco familiar trabajado en consanguinidad estrecha como base de nuestro criadero y varias ramas familiares paralelas en las que utilizaremos consanguinidad media-amplia. El uso de consanguinidad media-amplia nos permitirá conservar durante más tiempo la variedad genética de nuestro criadero y retrasará la aparición de los problemas derivados de la cría en consanguinidad.
En el momento que veamos que el grado de mejora se estanca será el momento de introducir un refresco para evitar la depresión consanguínea, con lo cual volvemos a empezar de nuevo, ya que introducimos un aporte genético que aumenta la variedad genética, disminuyendo el grado de homocigosis y alcanzando el llamado vigor de la heterocigosis; con la diferencia de que en la segunda generación no será preciso utilizar consanguinidad estrecha, pues la homogeneidad genética que habremos conseguido en nuestro criadero, si hemos hecho las cosas bien, hará indiferente emparejar los ejemplares de la primera generación del cruce de refresco con cualquiera de sus parientes.
Si trabajamos en equipo con otros criadores también podemos actuar de otra manera. Como hemos visto, a partir de la tercera generación es tal el número de combinaciones que podemos realizar que es imposible que un solo criador pueda hacerlas todas. Si trabajamos en equipo con otros criadores, podemos pasarles ejemplares de nuestro tronco familiar para que exploten esas opciones que por falta de espacio o de tiempo nosotros no podemos desarrollar. De esta manera, podemos encontrarnos con varios criadores trabajando un mismo tronco familiar (estirpe) que por efecto de la deriva genética y después de varias generaciones, conservarán las características principales del original tronco familiar pero con peculiaridades propias de cada criadero (ahí podremos hablar de varias líneas de la misma estirpe familiar).
Si el trabajo se hace de forma coordinada e inteligente, unidad de criterios a la hora de fijar los objetivos a conseguir y en la forma de seleccionar los reproductores y confeccionar las parejas, y además no se produce intercambio de ejemplares entre los integrantes del grupo durante varios años, cuando una de las líneas empiece a mostrar síntomas de declive podremos refrescarla con ejemplares procedentes de otra de las líneas paralelas, la consanguinidad será bastante amplia (posiblemente en este momento estemos hablando ya de parentescos de 6º grado en adelante) y aunque los patrones de emparejamientos hayan sido similares y hayamos buscado lo mismo, la deriva genética hará que se produzca el deseado efecto de recuperación de la variedad perdida y consigamos el mismo efecto que si introdujéramos un ejemplar no emparentado, con la ventaja de que en nuestro caso tendremos ganado bastante terreno y evitaremos tener que perder un par de años si los resultados del cruce de refresco no emparentado no son los deseados.
Puedo dar fe de que esto es posible y de que se hace con gran éxito en la actualidad por varios grupos de aficionados. Aunque en honor a la verdad hay que reconocer que el intercambio de ejemplares entre algunos criadores se hace antes de lo que sería deseable, perdiendo con ello posibilidades de aumentar la deriva genética recomendable en este sistema de desarrollo de una estirpe en varias líneas familiares paralelas.
5º) Aunque sigamos un esquema como el puesto como ejemplo en el punto anterior, debemos tener claro que no debemos ser prisioneros del mismo y que para hacerlo bueno debemos tener presente en todo momento las reglas 1ª, 2ª y 3ª. Cuando planifiquemos un emparejamiento siempre debemos preguntarnos primero por qué y para qué, por mucho que estemos siguiendo un esquema si el cruce no sirve al objetivo perseguido no se hace y punto; sobre todo cuando se trabaja en consanguinidad. La consanguinidad por sistema o porque sí es una soberana tontería, debe tener un fundamento.
6º) Cuando hagamos cruces abiertos hemos de procurar que los ejemplares provengan de familias de características afines y seleccionadas y depuradas mediante consanguinidad.
7º) Tengamos presente que no es lo mismo un sujeto campeón que un raceador, aunque un campeón puede ser un raceador los hay que, al proceder de cruces abiertos, no racean o dan una descendencia poco uniforme respecto a las características deseadas.
8º) No todos los ejemplares racean o transmiten igual las características deseadas, hemos de tener identificados aquellos ejemplares, machos o hembras, de nuestros criaderos cuyos descendientes demuestren en mayor medida las características deseadas y centrar nuestro trabajo en ellos. Si estudiamos los pedigrís de los mejores canarios, veremos en los mismos que hay ejemplares que se repiten una y otra vez, demostrando su condición de raceadores14. _____________________________________________________________________________
1 De Baseggio, Giorgio: "Campioni e Razzatori", segunda edición - Mondo degli Uccelli, Camugnano, 1996.
2 Para más información consultar la siguiente dirección de Internet: http://www.geocities.com/eltimbradoatc/articulos/eltriangulovida.html
3 Sobre el resto de factores recomiendo la lectura de: Martín Espada, Miguel Angel: "Consideraciones sobre la Herencia en los canarios de canto"; "Influencia de los meses de voladero en el canto del canario" y "Preparación de los canarios de canto para los concursos"; pueden consultarse en la sección artículos de la Web: www.cantodiscontinuo.com
4 Dalton, D.C.: "Introducción a la Genética Animal Práctica", Editorial Acribia, Zaragoza, 1982.
5 García Moreno, Valeriano: "Cría en Consanguinidad", Revista Pájaros nº 24, 3ª época, Madrid 1987.
6 Abellán Baños, José Antonio; "La quinta generación (R-4) del Canario Jaspe", Revista Pájaros nº 61, cuarto trimestre, Dos Hermanas 2004.
7 Drove Aza, Antonio: "No existe tal enigma…", Revista Pájaros nº 4, 1ª época, Madrid 1959.
8 Recomiendo la serie de artículos sobre canarios de Postura de Juan Moll Camps en la Revista Pájaros.
9 Buxade, Carlos y otros: "Zootecnia. Bases de Producción Animal", Tomo IV, Ediciones Mundi.Prensa, Madrid 1995
10 Se podrían utilizar diferentes clasificaciones combinables entre sí, como por ejemplo:
- Por el tipo de canto: doméstico, asilvestrado, etc.
- Por la especialización tímbrica: brillante, acuoso, hueco, etc.
- Por el tipo de giros fundamental predominante: de floreos, de floreos lentos, de variaciones conjuntas. 11 Ver nota 3.
12 El autor, junto a Santiago Iso Gracia, realizó un experimento consistente en cruzar canarios discontinuos con hembras de raza Gloster, para ver como se transmitía el patrón innato o genético de canto. El experimento confirmó las hipótesis de trabajo propuestas en este artículo.
13 Meiosis: doble división de las células germinales que reduce a la mitad el número de cromosomas y, por tanto, origina células haploides. La meiosis es un mecanismo adecuado para la distribución de genes entre los gametos, de tal forma que permite su recombinación y segregación al azar. Fuente: Enciclopedia Microsoft Encarta.
14 De la Coba Garrido, Alfonso: "Canto y Genética", puede consultarse en la sección artículos de www.cantodiscontinuo.com.