Concepto de raza: concepto y realidad
Publicado el
por Gabriela.
AUTOR: Miguel A. Martín Espada
Raza. Concepto científico
RESUMEN
Ante la controversia sobre el concepto de raza, agudizada por las noticias referentes a la igualdad del genoma humano y por las curiosas definiciones de la FAO respecto a las razas de ganado, que ha supuesto para muchos la negación o inexistencia de dicho concepto, se intenta defenderlo a partir de una serie de argumentos objetivos y científicos.
En primer lugar, resaltando el relativo valor de la aparente similitud genómica a partir del ADN nuclear observada en los humanos y, sin embargo, la gran importancia de otros factores, conocidos o no, entre los que destaca la influencia del medio externo e interno sobre la diferenciación entre individuos.
Por otra parte, se defiende que la formación de las razas no es un proceso moderno, sino por el contrario, muy antiguo y conocido históricamente aunque sin duda acelerado y ampliado en los últimos tiempos.
Igualmente se considera que la identificación y diferenciación de las razas se basa principalmente en criterios fenotípicos o externos (morfológicos, fanerópticos, fisiozootécnicos, etc.), mientras ciertos marcadores genéticos (microsatélites, etc.), por el momento, solo suponen un complemento o apoyo a esa diferenciación racial.
Otro aspecto que refuerza la existencia del concepto de raza, es precisamente el distinto comportamiento en cuanto a la transmisión de los caracteres, bien si se considera en el interior de la raza (selección) o entre razas (cruzamiento), aportando claros ejemplos a partir de razas originarias del mismo tronco.
Finalmente otro argumento para la defensa del concepto de raza viene dado por la moderna y seria actividad conservacionista de las razas en peligro (defensa de genes peculiares) y por el desarrollo científico de los numerosos esquemas de mejora genética realizados en todos los países y apoyados de forma oficial.
INTRODUCCIÓN
No es fácil en el momento actual hablar de etnología, de raza o de cualquier otro concepto que de alguna manera esté vinculado con dichas acepciones, sin levantar discusión o polémica.
Todo ello es debido a una cierta desorientación, incorrecta información, desconocimiento de la realidad ganadera o incluso, desgraciadamente, por interés malintencionado.
Por otra parte si no se cree en el concepto serio de raza, o se le aplica una valoración meramente ambiental, cultural o incluso folclórica, difícilmente se podrá considerar la necesidad de su conservación, ni tampoco serían consecuentes los numerosos esquemas científicos de mejora desarrollados en la actualidad para cada raza.
Vamos por ello a intentar defender con argumentos objetivos el concepto de raza y en buena lógica a la ciencia que la estudia (Etnología Zootécnica, en nuestro caso) para finalmente analizar el interés de la conservación y promoción de las mismas, todo ello a la vista de la evolución acaecida en estos últimos años.
CONCEPTO DE RAZA
La idea de raza ha recibido numerosas definiciones, pudiendo referirnos en este sentido a las documentadas revisiones de Rodero (1998) y Herrera (2001). Curiosamente su evolución ha sido notable desde aquéllas iniciales, rotundas y curiosas, pero a la vez inteligentes, como la del propio Darwin indicando que la raza, la variedad son especies nacientes o en vías de formación, definición que defiende claramente la existencia de dicho concepto.
Por otra parte numerosos etnólogoszootécnicos, buenos conocedores de la producción ganadera, mantienen definiciones concretas y claras, como las expuestas por algunos clásicos españoles. Así:
Colectividad de individuos que pos e en un conjunto de caracteres distintivos y transmisibles por generación (González Pizarro, 1903).
Conjunto de individuos con caracteres morfológicos, fisiológicos y psicológicos propios, por los que se les distingue de otros de su misma especie y que son transmisibles por herencia dentro de un margen de fluctuación conocido (Aparicio Sánchez, 1956).
Existen otras más modernas, en la misma línea, que incluso reconocen el hecho real y la importancia de las asociaciones de ganaderos y de los libros genealógicos:
Grupo de animales de características similares que reproduciéndose entre sí dan una progenie del mismo tipo, dentro de los estándares publicados por la organización de registro (Alderson, 1974).
Incluso el propio Cavalli-Sforza (2000) nos indica como definición de raza: Un grupo de individuos que pueden ser reconocidos como biológicamente diferentes respecto a otros. Definición concentrada y de alto valor científico, pero nada cultural o geográfica.
Recientemente en el volumen de la FAO, World Watch List for domestic animal diversity y en su tercera edición (Scherf, 2000), se observa una razonable, aunque incompleta, definición de raza que ha sembrado dudas: Grupo subespecífico de animales domésticos con características externas definidas e identificables que le permite ser diferenciado por apreciación visual de otros grupos definidos de la misma especie. Faltan aquí aspectos tan fundamentales como la transmisión a la descendencia de dichos caracteres, hecho de dinámica genética de gran importancia, no considerando tampoco caracteres no estimables por la simple visión, pero sí muy definitorios de una raza (crecimiento, producción lechera, etc.).
Además, junto a esta corta definición, añade otra posible variante como grupo de animales para el cual la separación geográfica y/o cultural de otros fenotípicamente similares, le ha permitido que se acepte para e l los una identidad distinta. ¿Entiende Scherf lo que quiere decir fenotípicamente?. ¡Qué criterios tan superficiales y escasos emplea la FAO para intentar definir el concepto de raza!. Para colmo añade un comentario, en la línea de todo lo anterior, que es verdaderamente poco afortunado: En este sentido la raza no es un término técnico. La raza es a menudo aceptada más como un concepto cultural que técnico.
Ante estas últimas ideas desearía aportar una definición personal para reafirmar y aclarar nuestra posición: Raza es un concepto técnico-científico, identificador y diferenciador de un grupo de animales, a través de una serie de características (morfológicas, productivas, psicológicas, de adaptación, etc.) que son transmisibles a la descendencia, manteniendo por otra parte una cierta variabilidad y dinámica evolutiva.
Otra circunstancia que complica la situación es la existencia de contrarios al reconocimiento de las razas como entidades taxonómicas, denominados etnoescépticos (Rodero, 1998), entre los que se incluirían a Lerner y Donald (1969), Blasco y Santacreu (1992) y Orozco (1995), que consideran las razas como producto exclusivo del hombre. Este hecho es en cierto modo explicable por la concreta especialización de algunos de ellos (avicultura y cunicultura), especulaciones que en la zootecnia moderna se mueven a partir de neo-razas, cruces, híbridos, e t c .
Por último las numerosas declaraciones que diferentes investigadores (Craig, Wallace, Freeman, etc.) han realizado recientemente en el sentido de las escasas diferencias genómicas encontradas entre las diversas razas de la especie humana, hablando incluso de eliminar este concepto, no han sido muy positivas y sobre todo han creado una notable desorientación por una parte y quizás un encastillamiento en los detractores de la etnología en general y de la zootécnica en particular.
Esta curiosa evolución del concepto de raza nos ha hecho meditar sobre el tema, por lo que creemos necesario y posible argumentar en su defensa de forma objetiva.
LAS RAZAS Y EL GENOMA
Es cierto que al realizar el estudio comparativo del genoma humano se han encontrado escasas diferencias inter-raciales y por ello algunos científicos han retendido invalidar el concepto diferencial de raza y su propia existencia. Existen dos tipos de razones que justifican estas conclusiones. De una parte, razones sociológicas. Así Cavalli-Sforza dice que no me gusta la palabra raza. Evidentemente porque él la utiliza en sus estudios sobre la especie humana y entonces le repugna en cierto modo su uso, ya que puede aparecer el fantasma del racismo, cosa que no desea. Pero no es porque el término conceptual de raza no se manifieste también en la humana condición y tenga una clara base biológica, es decir científica. La postura de Cavalli-Sforza y de otros muchos investigadores, entre ellos algunos del Human Genome Project, se basa en el pudor y en la prevención por circunstancias sociopolíticas y no científicas. Pero nosotros nos referimos a animales domésticos y por tanto esa prevención no tiene por qué existir, no repugnando utilizar el concepto raza de forma objetiva y seria en nuestro ganado.
Por otra parte existen razones científicas. Así, es necesario advertir que en los estudios del genoma humano, se ha utilizado como patrón base las secuencias de ADN del núcleo celular. Pero existen también otros factores de posible diferenciación no utilizados en el estudio, como el ADN mitocondrial o muy especialmente el propio medio, tanto externo como interno.
El externo al individuo (clima, nutrición, acción social, religiosa, aprendizaje, orientación selectiva, migraciones, patología ambiental, etc.) puede lógicamente diferenciar de forma determinante a un grupo de seres vivos a lo largo de los años en una concreta dirección, tanto en los casos en que haya existido cierta endogamia, como en aquéllos en los que se produjeron notables cambios de medio o cruces entre grupos de animales por efectos migratorios o dirigidos por el hombre.
Pero además el medio interno (diferencias en el sustrato celular, histológico, anatómico, enzimático, hormonal, interacciones posibles, etc.) también puede llegar a provocar respuestas totalmente distintas a partir de similares condicionantes genéticas iniciales, haciendo que seres de la misma especie, por efecto del medio (externo e interno), puedan evolucionar de forma muy diversa, conformando grupos con diferencias notorias entre sí, lo que perfectamente podría dar lugar a las razas.
Así es muy posible la existencia de otra serie de parámetros, desconocidos por ahora, que pudieran actuar como diferenciadores (los llamados marcadores específicos o diferenciadores raciales).
En definitiva, y de forma simplista, el genoma está constituido por un grupo no elevado de ladrillos diferentes, que son sin embargo acoplados de mil formas distintas, tilizando cemento, arena, agua, pintura, etc. muy variados, bajo la dirección de arquitectos y albañiles diversos, obteniéndose en consecuencia edificios (grupos animales o razas) muy distintos. Sin embargo la base analizada y comprobada es la misma (ladrillos), de ahí las escasas diferencias que entre razas muestra el genoma.
Por tanto la utilización de los resultados de la secuencia genómica entre las razas humanas (sólo el 0,01 p.100 de diferencia y en consecuencia el 99,99 p.100 de similitud), no sirven para concluir que no existen diferencias suficientemente claras, ya que solamente nos indican que sustancialmente los ladrillos empleados para construir el genoma humano son los mismos. Sin embargo todos pueden comprobar cómo las diferencias morfológicas, de coloración, etc. son bien diferentes entre las distintas razas
humanas y cómo incluso las aptitudes y orientaciones son evidentemente también diversas. Es el caso bien observado en atletismo, ya que incluso en el interior de las mismas agrupaciones étnicas melánicas son diferentes las morfologías y las aptitudes del tipo americano (velocidad y salto), respecto al africano de las altas mesetas (carreras de fondo).
En las razas de ganado, las diferencias en las secuencias de ADN estudiadas son también pequeñas, no sólo entre razas, sino incluso entre especies distintas.
En este sentido las diferencias genómicas entre una vaca y una oveja son también enormemente pequeñas. Sin embargo no sólo son razas distintas, sino claramente aceptadas como especies tan diferentes, que incluso no pueden reproducirse entre sí.
Efectivamente (De Blas y Fraga, 1988 y Wolf, 1990) si comparamos la hormona de crecimiento bovino (BST) con la ovina (OST) observamos una escasa diferencia en la secuencia de ADN (sólo un aminoácido distinto entre 190, es decir el 99,5 p.100 de igualdad). Son especies bien diferentes pero sus hormonas de crecimiento son muy similares, de forma que la BST puede ser empleada sobre la oveja (Requena, 1996) sin rechazo de ningún tipo y con positiva respuesta (incremento notable de la producción lechera y de su crecimiento). ¿Son por ello especies iguales?. Evidentemente no, a pesar de la similitud de su ADN.
Entre la hormona de crecimiento porcina (PST) y la de la rata (RST) sólo existen 10 aminoácidos diferentes (94,7 p.100 de igualdad). Sin embargo, no podemos atrevernos a decir que el cerdo y la rata sean especies similares a pesar de que inoculando PST en diferentes animales de laboratorio se obtienen mejoras en el crecimiento (Knobil y Greep, 1959).
Siguiendo el mismo razonamiento, las diferencias globales a nivel de la secuencia genómica entre un hombre o un ratón, también son pequeñas, pues son animales vertebrados, de sangre caliente y mamíferos, lo que condiciona grandemente esa similitud, sin embargo está claro se trata de especies enormemente distantes y distintas.
No parece por tanto muy válido el método de intentar diferenciar razas intra especie a partir de la secuencia genómica del ADN nuclear, al menos a la luz de los conocimientos actuales, ya que el sistema no es suficientemente válido ni siquiera para diferenciar especies próximas.
Las numerosas combinaciones de adenina, timina, citosina y guanina (ATCG) en la doble hélice del ADN no explican claramente la funcionalidad de esa variada disposición, que sobre la base de interacciones diversas medioambientales o a la diferente naturaleza del soporte correspondiente, pueden provocar respuestas bien distintas, como dijimos.
El conocimiento actual no nos faculta todavía a establecer esa simplista proporcionalidad (causa-efecto) entre secuencia de ADN y respuesta o carácter fenotípico posible. En una palabra, y como ejemplo simple, incluso pueden existir genes detectables, que sin embargo nunca serán activados, es decir no son operativos, no actúan, o sólo en determinadas condiciones ambientales, lo que puede falsear los resultados de diferenciación e identificación.
Estos hechos, desconocidos por ahora en cada caso, son los que evidentemente pueden marcar notorias diferencias entre genomas aparentemente muy similares, siendo quizás los posibles causantes de esa diversidad aún no explicada.
En definitiva queda todavía mucho por hacer y conocer, como ya dijimos. Nos hallamos iniciando el camino de la proteómica, ciencia que estudia los procesos que se producen en el interior del genoma y que en su momento podrían explicarnos más y mejor por qué seres vivos tan teóricamente similares, son realmente tan diferentes, explicando y confirmando así de forma científica el concepto de raza.
Por otra parte si los genomas fueran tan iguales, en estructura y funcionalidad, no hubiera sido posible alcanzar la evolución tan determinante que han sufrido los seres vivos. En una palabra, negar el concepto de raza o variedad sería negar su proceso evolutivo, o la propia deriva biológica de los mismos (evolución o deriva naturales o
conducida por el hombre), ya que no puede todo dejarse en manos de la mutación.
Quizás la estructura del ADN pueda ser razonablemente parecida, pero posiblemente la funcionalidad sea muy diferente.
A veces pensamos si no se tratará más bien de una mera discusión semántica y no científica, que gira entorno al hecho de adjudicar o no a este concepto el apelativo de raza, lo que para algunos parece denostable por motivos poco claros, posiblemente sociales y políticos, pensando especialmente en los humanos, como ya hemos dicho anteriormente.
El concepto y el hecho diferenciador de diversos grupos animales dentro de la misma especie seguirá existiendo, aunque podamos llamar a estos subconjuntos, dentro del conjunto especie, de la forma que queramos acordar (raza, subespecie, variedad, etc.).
CREACION DE LAS RAZAS
Algunos autores afirman que la creación real de las razas de ganado es muy moderna, ofreciendo una antigüedad de sólo unos 200 años, coincidiendo con la acción de los criadores ingleses (Bakewell, Ellman, hermanos Colling, etc.) y de otros europeos en los siglos XVIII y XIX. Sin embargo cuando los ganaderos ingleses iniciaron su labor ya existían en sus campos razas bien definidas, tanto de vacuno (Shorthorn, Devon, Hereford, Highland, etc.) como de ovino (Ryeland , Cotswolds, Lincoln, etc.).
En consecuencia son hechos muy anteriores al siglo XVIII tanto la creación de las razas como su posterior evolución, pues se inicia ya con la domesticación, en línea con las ideas de Denis (1982) y lo expuesto por Rodero (1998). Antes de la domesticación únicamente actuaba la selección natural. Posteriormente la acción consciente y directa del hombre a través de una cierta reproducción dirigida en base a una selección fenotípica o masal sobre determinados caracteres morfológicos, productivos, etc. o incluso por cruzamientos con otros grupos animales debido a migraciones o por elección humana, todos ellos hechos genéticos concluyentes desarrollados de forma
deliberada por el hombre o de manera aleatoria.
Todo esto, unido a las particulares condiciones medioambientales que rodeaban a cada una de las ya inicialmente caracterizadas poblaciones animales, fue singularizando y diferenciando grupos diversos de cada especie en el mundo, en cada país o región, conduciendo a la aparición y consolidación de razas concretas. Es cierto que al llegar el siglo XVIII estas acciones se potenciaron y desarrollaron de manera más ordenada y rigurosa, pero evidentemente ya existía una serie de razas bien diferenciadas e incluso muy reconocidas y demandadas. Igualmente se encontraban otras numerosas de tipo rústico o medioambiental, que han seguido evolucionando y perfeccionándose hasta nuestros días, como ya hemos indicado. Tomando como ejemplo la especie ovina y España, podemos resaltar la amplia iconografía existente en la Merina desde el siglo XV, mostrándose, ya en aquella época, perfectamente consolidada dicha raza (Aparicio y Pizarro, 2000).
Igualmente en el antiguo Reino de Aragón y en los documentos de la Casa de Ganaderos de Zaragoza, se describen claramente los tres tipos ovinos existentes ya en el siglo XV: Churro en la zona pirenaica (actual Churra Tensina), Merina en Teruel, especialmente en la serranía de Albarracín, aunque diseminada por todo Aragón, y la entrefina en el resto (Fernández Otal, 1996). De forma más concreta para la Rasa Aragonesa se dispone también de iconografía medieval y renacentista tanto en escultura (retablo y trascoro de La Seo, siglos XV y XVI), como en pintura (Cuadro de la Casa de Ganaderos de Tauste, siglo XVI) entre muchos otros, en donde se observa erfectamente dicha raza, blanca y mocha, con cabeza y patas deslanadas, subconvexilínea y de vellón entrefino.
Ejemplos como los citados podrían presentarse referidos a multitud de razas en todas las especies ya conformadas mucho antes de que los criadores ingleses apareciesen (cerdo ibérico, mastines mesteños, caballo árabe, etc.). ¿O es que vamos a pensar que no había ganadería y grupos animales perfectamente diferenciados en otros países?. Todo esto evidentemente no es contrario a que estos grupos animales hayan seguido evolucionando en el transcurso de los tiempos o incluso desaparecido, como clara cualidad inherente a cualquier raza.
Ejemplo claro de este proceso evolutivo son las primitivas razas autóctonas alemanas (rasseland) a partir de las cuales se formaron a lo largo del XIX y XX las veredelte rasseland o razas del país mejoradas.
En cualquier caso vemos cómo la construcción de una raza ha partido de un plan desarrollado por selección o cruce más o menos complejo, que unido a unos claros efectos ambientales ha determinado en conjunto la evolución y diferenciación de diversos grupos animales.
Resumiendo de forma esquemática el proceso de creación de las razas podríamos indicar:
1. BASE ANIMAL INICIAL Y GENÉRICA
- Creación y diferenciación de especies mediante evolución a largo plazo (selección natural, mutaciones, etc.) .
- Fuerte incidencia del medio y el azar.
2. ESPECIES ANIMALES INICIALES
- Evolución de cada especie:
a) Acción medioambiental inicial (hasta X-VII milenio a.J.C.)
a.1.- Selección libre, condicionada por el medio natural.
a.2.- Posibles influencias genéticas (mutaciones).
a.3.- Medio natural (clima, vegetación, orografía, etc.). Influencia geográfica, pero todavía no antropológica.
a.4.- Como consecuencia construcción inicial de grupos animales (subespecies, etc.) con características comunes.
b) Acción del hombre-domesticación (hasta II-I milenio a.J.C.)
b.1.- Comienzo de la reproducción dirigida. Iniciación a la selección y
cruzamiento.
b.2.- Influencia del medio más o menos modificado por el hombre.
c) Práctica de la reproducción dirigida (hasta el siglo XVII d. J.C.) Formación de grupos animales similares dentro de la especie (razas).
c.1.- Consolidación de la selección dirigida de tipo empírico pero objetiva y real (selección masal o fenotípica en función de caracteres morfológicos o productivos de interés para el hombre). Utilización general del cruzamiento. Etapa de enormes progresos en la diferenciación racial y en los resultados de mejora.
c.2.- Metodología genética muy simple y concreta, aunque a veces equivocada o mediatizada por otras circunstancias (ambiente, hechos culturales, religiosos, endogamia, errores de estimación, etc.), provocando un determinado sentido de evolución o de deriva del grupo animal correspondiente.
c.3.- Diferenciación. Como consecuencia de esta reproducción dirigida por el hombre se asiste a un rápido proceso de diferenciación animal por grupos, dentro de cada especie, formándose y consolidándose un elevado número de razas en un tiempo no exagerado. Basta leer a los autores antiguos para conocer las descripciones de las diferentes razas ya existentes y cómo los propios ganaderos realizaban no sólo selección, sino también cruzamientos para obtener más beneficio (carne, trabajo, lana, etc.). Recordemos a los clásicos Columela y Virgilio, a los autores árabes en el medievo o a la mayoría de los tratadistas europeos en la alta edad media y en el renacimiento.
Estas actuaciones no tenían nada de folclóricas o culturales, sino que estaban basadas en actividades serias, reguladas, objetivas, técnicas y concretas, con los lógicos errores debido a la escasa información y a los insuficientes medios de aquella época. Eran acciones directas de tipo genético (selección masal y cruzamientos), pensadas y realizadas por el hombre para modificar la base genética de un grupo animal, buscando unos fines fundamentalmente productivos y de adaptación, en definitiva económicos. Evidentemente el medio jugó un papel doble, ya que el hombre debía acomodar su incipiente esquema de mejora a las condiciones ambientales y a la vez el medio actuaba positiva o negativamente sobre lo que el hombre iba alcanzando o intentaba conseguir.
Así la acción geográfica (es decir, acción ambiental), lógicamente influía en tanto se derivaban de ella unas condiciones del hábitat (temperatura, humedad, suelo, orografía, vegetación, etc.) que habían ya ahormado las características iniciales de los grupos animales a partir de la selección natural (razas primitivas de Denis, 1982).
d) Acción moderna. Ganaderos ingleses y asimilados europeos y americanos (siglos XVIII y XIX)
Consolidación y mejora de numerosas razas ya existentes y creación de otras nuevas siempre en base a criterios productivos, mediante selección organizada, consanguinidad, amén de la utilización del cruzamiento.
Esto condujo paralelamente a la creación de asociaciones de las diferentes razas ya constituidas, lo que supuso una positiva colaboración por parte de los ganaderos facilitando así el citado proceso de consolidación, diferenciación y mejora de las razas ya existentes.
Evidentemente se desarrollaron a la vez una serie de hechos de tipo cultural y sociológico (exposiciones de ganado, concursos, premios, etc.) necesarios para mantener el apoyo de los criadores, lo que favorecía el afianzamiento de las razas.
En resumen, creación o mejora de una serie de razas con buen nivel productivo, mediante medios científicotécnicos objetivos y serios, ya alejados de la tradicional rutina, sentando las bases para los futuros esquemas de selección y cruzamiento.
Por otra parte múltiples razas ubicadas en países poco desarrollados, de gran rusticidad y productividad no elevada, pero ya perfectamente diferenciadas, se mantenían en procesos de consolidación y mejora todavía rutinarios (apartados b y c).
e) Acción contemporánea (siglo XX)
En los países desarrollados, y prácticamente en casi la totalidad del planeta, el siglo XX permitió la completa consolidación, diferenciación y mejora de muy numerosas razas, mediante la utilización de medios modernos con el apoyo de las estructuras ya iniciadas (asociaciones de ganaderos y libros genealógicos y de control de rendimientos). Constituyó una etapa en la que incluso fue posible crear, fijar y diferenciar diversas razas nuevas (synthetic breed), por supuesto mediante metodologías bien alejadas de criterios meramente culturales, por el contrario puramente científicos y buscando objetivos realistas y concretos (mejora de la prolificidad, capacidad de adaptación, producción de carne, leche, huevos, etc.). No olvidemos en este sentido que la popular Corriedale es en definitiva una de las primeras razas sintéticas de ganado creadas en el mundo en la época moderna. Así a finales del XIX se cruzó sabiamente Merina con Lincoln, añadiendo unas gotas de Leicester. Se intentaba por una parte conseguir una raza válida para zonas más húmedas (vellón más largo y abierto por acción del Lincoln), pero manteniendo una finura estimable (Merino) y con un vellón pesado, que ofreciera buen rendimiento económico.
Otras muchas se crearon con anterioridad mediante el tradicional sistema genético de cruzamiento, eligiendo los parentales adecuados para el objetivo previsto, y sometiendo posteriormente a selección y fijación la población obtenida (recordemos a Columela en su descripción sobre la formación de la raza merina).
Por otra parte ¿alguien puede discutir que los ovinos Assaf constituyen una diferenciada y reconocida raza lechera o piensan que es algo folclórico y cultural?.
En este sentido le preguntaríamos a los peculiares tratadistas de la FAO si consideran que todo lo anterior es meramente sociológico y cultural. Nosotros creemos que es algo distinto, más serio y que entra de lleno en lo científico, técnico y económico.
A veces algunos no comprenden claramente el concepto de raza y su importancia a la hora de poner en marcha un modelo productivo válido. Su desconocimiento de la realidad ganadera les supone un enorme lastre, hallándose, por desgracia, encerrados en singulares torres de marfil. Otros autores tienen una orientación concreta hacia la ganadería industrial en donde el mundo productivo se mueve especialmente a base de estirpes o híbridos comerciales. Sin embargo existieron y existen desde hace años y aún ahora una gran multitud de razas avícolas, cunícolas y porcinas, algunas de las cuales, por sus valiosas prestaciones, han servido de base precisamente para la creación de esos híbridos comerciales.
Esperemos que estas reflexiones, y las siguientes, ayuden a comprender mejor dicho concepto y su evolución en estos últimos años.
LOS CRITERIOS EXTERIORISTAS COMO BASE DEL CONCEPTO DIFERENCIADOR DE RAZA
Dos grupos de caracteres exterioristas nos permiten describir y diferenciar una raza (Sierra, 2001):
1.- MORFOLÓGICOS Y FANERÓPTICOS
Nos concretan determinado tipo de caracteres como el tamaño, peso, perfil, medidas, color, tipo de piel, pelo, lana, cuernos, plumas, etc. que en cierto modo suponen la marca de fábrica, permitiendo una rápida descripción y diferenciación racial de visu.
2.- CARACTERES FISIOZOOTÉCNICOS, EXPRESADOS FENOTÍPICAMENTE
Actúan como descriptores externos de una serie de parámetros fisiozootécnicos (reproductivos, productivos, de comportamiento o psicológicos, capacidad de adaptación, etc.), siendo estimados a partir de los registros o controles pertinentes.
En ambos casos existe una concreta base genética que sustenta dichos caracteres, por supuesto afectados por el medio, precisando tanto la serie de controles y medidas a realizar para su control, como su análisis y valoración posterior, una clara metodología científica.
El estudio coordinado de estos dos grupos de parámetros permite, en consecuencia, una seria y objetiva caracterización y diferenciación raciales.
3.- MARCADORES GENÉTICOS COMO DESCRIPTORES RACIALES
En principio, y hasta el momento actual, este grupo de marcadores, especialmente los microsatélites, no posee todavía una capacidad definitiva a la hora de describir y diferenciar razas de ganado. Sin embargo supone un apoyo complementario de gran importancia. Una interesante información referida especialmente al papel de los microsatélites en la diferenciación racial la podemos encontrar en Molina (2001).
No obstante, para algunos, la valoración de distancias genéticas entre poblaciones animales podría permitir diferenciar dichos grupos (razas) en virtud de las frecuencias alélicas correspondientes, ya que estas distancias pueden ser debidas tanto a mutaciones y deriva genética, como a selección natural y artificial.
Desgraciadamente los loci responsables de las diferencias debidas a dicha selección natural y artificial no suelen estar disponibles para el análisis de las distancias genéticas (Molina, 2001), con lo que difícilmente este método puede ser suficientemente fiable para diferenciar razas. Por el contrario el estudio de las distancias genéticas a partir de microsatélites puede ser más útil para evaluar la variabilidad intraraza (Barkeretal., 1993).
En definitiva para llegar a caracterizar y diferenciar mejor las razas sería preciso incluir en el estudio los caracteres morfológicos y fisiozootécnicos peculiares de la población problema (Ponzoni, 1997), tal y como ya hemos indicado con anterioridad.
Sin embargo otros autores (CavalliSforza et al., 1995), a pesar de sus prevenciones sociales sobre el término raza, aprovechan sus numerosos estudios sobre distancias genéticas para aproximarnos en la propia especie humana hacia una diferenciación entre grupos o razas. Así asumiendo que la distancia genética entre un inglés medio y un danés es de 1,0, encuentran que aumenta a 2,5 respecto a un italiano, se eleva a 9 frente a un iraní, multiplicándose la distancia 59 veces ante un japonés y hasta 109 si comparamos el inglés con un bantú.
Como vemos nuevas pruebas de argumentación genética (científicas por tanto) que rodean al concepto de raza.
EL CONCEPTO DE RAZA Y EL DESARROLLO DE ESQUEMAS DE MEJORA GENÉTICA
1- LA DIFERENTE TRANSMISIÓN DE LOS CARACTERES EN RAZA PURA O EN CRUZAMIENTO
Cuando se lleva a cabo un plan de mejora genética por selección sobre una determinada raza es obligado conocer las características de dicha raza y necesariamente su grado de pureza. Si no conocemos realmente el material con el que estamos trabajando podemos llegar a encontrarnos con curiosos y sorprendentes resultados (h2 de 0,3 a 0,5 en el carácter prolificidad en alguna raza ovina). La utilización de poblaciones comerciales, en las que conviven animales de una raza autóctona en pureza, junto con otros cruzados con múltiples razas, pueden explicar dichos resultados (Sierra, 2001).
Efectivamente en el desarrollo de un esquema de mejora por selección en raza pura, los criterios genéticos fundamentales que lo presiden son la heredabilidad y la repetibilidad del carácter o caracteres a mejorar. Por el contrario en un plan de mejora por cruzamiento (entre razas) la transmisión de los caracteres a la descendencia ofrece mecanismos bien diferentes. Así fenómenos de aditividad (determinismo genético aditivo) y de heterosis, provocan una mayor capacidad de transmisión en cualquier carácter.
Por eso se encontró una elevada h2 en la prolificidad en el caso antes citado, ya que no se estaba realizando un plan de mejora por selección intraraza, sino a partir de un colectivo de animales puros y cruzados en menor o mayor nivel, en donde los verdaderos criterios genéticos que presidían el esquema proporcionaban una variada y a veces mayor capacidad de transmisión de los caracteres frente a la esperable en raza pura. Se aplicó la fórmula para el cálculo de la h2 y los resultados fueron lógicamente muy superiores.
Este es uno de los hechos comprobatorios más importantes y contundentes que nos habla de la realidad de la existencia científica de las razas, ya que el comportamiento en cuanto a la capacidad de transmisión de los caracteres es totalmente distinto si se trabaja en el interior de una raza o si se realiza cruce entre razas. Por tanto, la existencia de las razas no está apoyada por razones de tipo cultural o sociológico, sino por argumentos genéticos, serios y objetivos, que marcan diferencias claras entre razas distintas.
2- DIFERENCIACIÓN RACIAL DE TRONCOS COMUNES
Tomando como base lo apuntado en el apartado anterior vamos a reflexionar a partir de los resultados de un grupo de razas originarias del mismo tronco y que mantienen por ello una razonable semejanza externa (morfológica y faneróptica), pero que sin embargo presentan caracteres fisiozootécnicos bien diferenciados.
Este es el caso por ejemplo de las razas Lacaune , Manchega y Rasa Aragonesa (Quittet, 1965, Esteban y Tejón, 1985 y Sierra, 1987). Animales blancos, mochos, de perfil subconvexo, lana entrefina, extremidades alargadas y desnudas y tamaño entre eumétrico y subhipermétrico. Su origen común (Ovis aries ligeriensis), en zonas próximas al macizo central francés y su periplo atravesando los Pirineos y asentándose en diferentes áreas (Aragón y La Mancha, incluso hasta Jaén y serranías de Granada, originando allí la Segureña), propició en cada zona la aparición de poblaciones animales que fueron evolucionando y diferenciándose en función del medio y de la selección dirigida que el hombre ejerció en cada lugar.
Así se originaron razas, perfectamente aceptadas como tales, que se diferencian en la actualidad especialmente por sus características fisiozootécnicas (crecimiento, prolificidad o producción de leche), manteniendo un aspecto externo bastante similar.
El hombre, en base a las posibilidades del medio y a sus objetivos particulares, se preocupó de la mejora productiva en unos casos, ejerciendo por el contrario en otros una labor selectiva poco relevante.
Así los esfuerzos en mejora de leche y prolificidad de la raza Lacaune son bien conocidos, manteniendo esquemas de mejora que han permitido magníficos resultados en la actualidad (Barillet, 1985). En el caso de la Manchega ha existido siempre una vocación hacia el ordeño, aunque los trabajos serios en mejora de la producción lechera son todavía recientes (Pérez Guzmán et al., 1996). Finalmente la raza Rasa Aragonesa, en virtud del difícil medio que siempre la ha rodeado, se ha mantenido como una raza ambiental y rústica, sufriendo incluso durante años una selección negativa en el caso de la prolificidad (1,08 a 1,20 según época de cubrición en los años sesenta, Sierra, 1972).
Esto nos ofrece un ejemplo claro de dinámica evolutiva diferente en una serie de grupos raciales de origen común a causa del medio y de la selección dirigida, dinámica evolutiva que ha desembocado en la creación de razas bien distintas.
En este sentido las diferencias genéticas existentes entre estas tres razas se hallan consolidadas, siendo perfectamente demostrables, ya que su comportamiento genético en selección intra-raza y en cruzamiento lo avalan (tabla I).
Así el progreso genético por selección de la producción lechera en la Manchega es lógicamente lento (h2: 0,25-0,30), sin embargo el simple cruce con sementales de raza Lacaune ha permitido en las hembras cruzadas F1 cifras de alrededor de 180 litros por lactación en virtud del determinismo genético aditivo, que posibilita en la F1 registros iguales a la semisuma de las producciones de las razas parentales (175 litros teóricos), añadiendo a ello del 3 al 5 p.100 por heterosis.
Lo mismo ocurre en el caso de la prolificidad, carácter de muy difícil mejora en raza pura debido a su baja h2 (0,03-0,10).
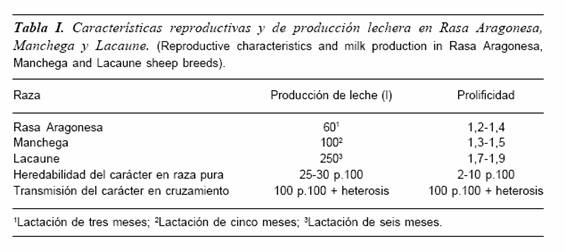
En este mismo cruce entre Lacaune y Manchega las F1 en segundo parto han ofrecido resultados de 1,5-1,7 corderos (Sierra, 2000).
En definitiva se ha producido una evolución genética diferenciadora a partir del mismo tronco originario común, en función del medio y de la acción del hombre, provocando la creación y fijación de razas diferenciadas, como lo prueban no sólo sus registros productivos distintos, a pesar de su aspecto externo bastante similar, sino especialmente sus diferentes comportamientos genéticos (selección intraraza vs cruzamiento entre sí) en la transmisión de los caracteres.
De ahí se derivan precisamente una serie de prácticas comunes de gran alcance genético que permiten una mejora más rápida utilizando diferentes tipos de cruzamientos entre razas distintas. Así, el empleo de razas como líneas padre (sire line) sobre otras razas (líneas madre o dam line), metodología científica de tanta trascendencia en la mejora de carne, leche, prolificidad, etc. en todas las especies ganaderas o la creación de razas sintéticas a partir del empleo de razas diversas.
De nuevo argumentos científicos objetivos y serios que avalan el concepto de raza, muy lejos de los culturales y sociales esgrimidos por la FAO.
CONSERVACIÓN Y ESTUDIO DE LAS RAZAS DE GANADO
Otro de los argumentos científicos fundamentales a la hora de defender el concepto de raza es precisamente la moderna actividad conservacionista que sobre dichos grupos de animales se está desarrollando.
Si las razas en realidad no existieran como tales o sólo fueran colectivos de animales diferenciados únicamente por criterios de tipo histórico, cultural, etc., o bien estuviesen reconocidos como agradecimiento al apoyo que durante siglos ofrecieron al hombre
(alimento, vestido, trabajo, etc.) o como antiguos monumentos biológicos que es preciso mantener (Sierra, 1994), sería comprensible una acción de conservacionismo basada en criterios históricos, culturales o de agradecimiento.
Sin embargo los esfuerzos que numerosas organizaciones y asociaciones tanto nacionales, como internacionales (Rare Breed International, EAAP, FAO, UE, etc.) están realizando en pro de la conservación de las razas, se deben fundamentalmente a la preservación de la biodiversidad doméstica, evitando la posible pérdida de genes valiosos como consecuencia de la desaparición de dichas razas. Estos genes suponen un caudal precioso per se, que además podría ser reutilizable en un futuro próximo, transmitiéndolos a otras razas, quizás más productivas pero con deficiencias que podrían limitar su rentabilidad (rusticidad, resistencia a determinadas enfermedades, fertilidad, calidad de carne, etc.).
Por tanto uno de los objetivos fundamentales del conservacionismo de las razas es claramente científico y no sólo cultural o de agradecimiento. Otro argumento más para defender el concepto de raza desde la propia etnología zootécnica y la genética.
Numerosos autores han sentado las bases de dicho conservacionismo científico en las razas del ganado (Brem et al., 1984; Maijala, 1990; Alderson, 1990; Bodo, 1990; Simon y Buchenauer, 1993, etc.), ordenando y regulando los mecanismos de conservación a través de reuniones, congresos y publicaciones específicas en el entorno de la FAO (1990), a lo que se unen sus ediciones de la World Watch List for Domestic Animal Diversity, junto con la tradicional revista Animal Genetics Resources.
En este sentido es preciso recordar la labor realizada en España desde 1986. Primero a través de la creación de la Sociedad Española pro Recursos Genéticos Animales (SERGA) (Sierra, 1987) cuyo objetivo básico es la conservación de las razas en peligro, con una amplia actividad desarrollada desde su comienzo, asistiendo a las iniciales reuniones comunitarias a fin de preparar la primera lista de razas de rumiantes en dificultades a petición de la Comunidad Europea, que permitió sentar las bases para la concesión de subvenciones a las mismas y el aporte de interesante información para el Genetic Data Bank de Hannover.
Así, numerosas actividades fueron consolidando el buen hacer de SERGA, desde su reconocimiento internacional al incorporarse a la RBI (Rare Breed International) en 1992, pasando por la asistencia a numerosos congresos y reuniones internacionales sobre el tema, colaborando en la recogida de información de nuestras razas en peligro en todas las especies como punto focal para los fondos internacionales de la Unión Europea, EAAP, FAO, etc. y participando en la red de información mundial DAD-IS. Igualmente en el plano nacional su labor de colaboración científico-técnica con el MAPA, cabildos y diputaciones de las diferentes autonomías, centros de estudio e investigación, etc., están suponiendo el desarrollo de proyectos y asesorías muy diversos sobre conservacionismo ganadero.
Los varios congresos científicos nacionales e internacionales organizados por SERGA y la publicación de su revista El Arca en 1997, han sido igualmente reflejo de una actividad seria e imparable. Actividad que tendría menos razón de ser si las razas sólo ofrecieran un interés cultural, geográfico, etc . Pero evidentemente el campo de la Etnología es más amplio que el referido a las razas de escaso censo.
Por ello, y desde hace unos años, algunos estudiosos de la Etnología zootécnica pensaron en la necesidad de llevar a cabo una puesta al día de sus objetivos, ofrecerlos al público científico con una nueva filosofía que resaltara su verdadera importancia y por supuesto que amparase en su estudio y preocupación absolutamente a todas las razas de nuestras especies domésticas.
En este sentido un grupo de etnólogos celebraron el I Encuentro de Docentes e Investigadores Zooetnólogos españoles (SEZ, Córdoba, 2001) en donde fue creada la Sociedad Española de Zooetnología. Esta sociedad, de tan joven creación, ha iniciado ya sus actividades como colaboradora del MAPA en todo lo referente a nuestras razas ganaderas y desde aquí, pues participamos de su ilusión y objetivos, le deseamos el más próspero futuro.
Por último en todos los países desarrollados los ministerios correspondientes, en general de Agricultura, realizan inventario y clasificación de sus diversas razas, describiéndolas a través de catálogos oficiales y apoyándolas mediante concursos, subastas subvencionadas, etc. y manteniendo los correspondientes comités oficiales de razas de ganado.
Igualmente se hallan establecidas numerosas asociaciones de ganaderos de las diferentes razas que desarrollan programas científicos de mejora genética a partir de sus libros Genealógicos y de Control de Rendimientos, siendo incluso dichas razas soporte en algunas casos de productos de calidad (queso, jamón, carnes frescas, etc.) reconocidos oficialmente, no sólo en cada estado, sino por la propia Unión Europea.
Nuevas y sólidas aportaciones para defender de forma objetiva, científica y técnica el concepto de raza, que no puede cobijarse bajo el simple epígrafe de geográfico, cultural o folclórico.
Sierra Alfranca, Isidro
Catedrático de Etnología y Producción Animal. Facultad de Veterinaria. Universidad de Zaragoza.