Aprendizaje del canto en las aves
Publicado el
por Gabriela.
AUTOR: Gilberto Haddad
APRENDIZAJE DEL CANTO EN LAS AVES, DIVERSIDAD Y PLASTICIDAD, OPORTUNIDAD Y RETOS
El sistema neuronal que regula el aprendizaje del canto en las aves cantoras, se ha convertido en un modelo prominente para el estudio del mecanismo neuronal de aprendizaje. En varias especies de aves, el aprendizaje del canto es caracterizado por un periodo sensitivo bien definido. También se ha observado que el aprendizaje del canto y su producción son controlados por un discreto y bien definido circuito neuronal. El canto es esencial para el comportamiento reproductivo de las aves y provee una oportunidad de estudio del sistema neuronal básico del comportamiento de un aprendizaje en un contexto naturista. Hay más de 4000 especies de aves cantoras, las cuales muestran una extensiva diversidad en diferentes aspectos del aprendizaje del canto. (Brenowitz y Beecher, 2005).
Proyección del Núcleo Mayor en el Sistema De Control de Canto
Las vías motoras (verdes) controlan la producción del canto y consisten en el descenso del HVC (Acronym) del Nidopallium al Núcleo Robust, núcleo del Arcopallium (RA), para luego dirigirse del núcleo vocal (Nxllts) “parte traqueo siringeal del núcleo hipogloso”, al núcleo respiratorio Retroanbigualis (RAM) y al núcleo Ambiguo Laringeal (AM) en la medula. Las neuronas motoras (Nxllts) inervan los músculos de la siringe, el órgano de producción vocal de las aves. Las líneas azules indican los fascículos aferentes del HVC del núcleo talamico Uvaeformis (UVA) y del núcleo interfase Nidopallial (NLF). Las líneas rojas indican los fascículos auditivos que van al NFL y al HVC de la región telencefálica de la audición. Las líneas anaranjadas indican las vías cerebrales anteriores (AFP), la cual es esencial para el aprendizaje del canto y la percepción. Conectando en forma directa al HVC con el RA, vía área “X” (Ganglio basal homologo), la porción medial del núcleo dorso lateral del tálamo (DLM) y la porción lateral del Núcleo Magno Celular del Nidopallium (IMAN). El IMAN solo proyecta al área “X”, además de una línea adicional hacia los ventrículos (V). (Brenowitz y Beecher, 2005).
Periodo Sensitivo Para Memorización Del Canto
En este periodo de aprendizaje de canto existen cambios correlativos en la estructura y fisiología del sistema de control de canto. Existe una fase sensitiva, la cual es esencial para el normal aprendizaje del canto.
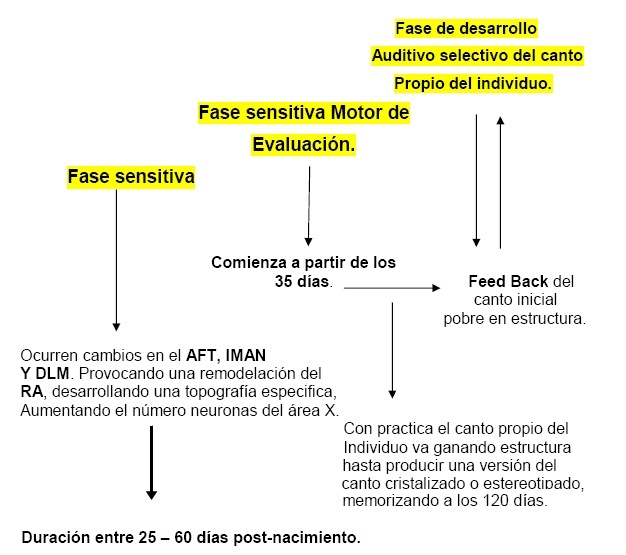
Uno de los grandes enigmas al estudiar estas áreas de aprendizaje, es referido a que si estos cambios anatómicos y fisiológicos, ocurren como procesos normales del desarrollo del individuo o están activamente correlacionados con el aprendizaje del canto. Es por ello que el desarrollo del sistema de canto esta todavía hoy en día en estudio, observando que los cambios celulares podrían estar relacionados a eventos desarrollados independientes al proceso de aprendizaje del canto.
¿Serán estos cambios anatómicos funcionalmente relacionados al aprendizaje o son parte de un proceso de desarrollo general?
Esta respuesta se podría responder con el estudio de aves (Passerina cyanea) con memorización y ensayo de canto retardado, hasta el final del primer año o principio del segundo o en especies de memorización y nuevo ensayo de canto cuando adultos (Starlings sp y Serinus canarius). Donde se podría predecir en el primer grupo de aves, de aprendizaje tardío, si los cambios anatómicos están relacionados realmente con el aprendizaje del canto. Observando los cambios tardíos que ocurren en el circuito de canto. En aves que memorizan nuevos cantos cuando adultos, se podría observar si un patrón juvenil de conexiones sinápticas, conseguirían ser restauradas o mantenidas, dependiendo si el nuevo canto es memorizado anualmente o continuamente.
Si el LTP y el IMAN, estarían relacionados con la memorización, podríamos predecir un retardo en el movimiento de potenciación o depresión sináptica en especies con aprendizaje diferido (Song sparrows y Índigo Bunting) y una restauración o mantenimiento del LPT en especies con nuevo aprendizaje de canto cuando adulto (Serinus canarius). Esto nos permite inferir que aunque la neuro-génesis juega un papel importantísimo en el aprendizaje del canto, todavía no se tiene la información necesaria y completa para relacionar estos dos procesos en las diferentes especies de aves de canto.
Doupe (2004) demostró que existen cambios en la AFT en el periodo de aprendizaje de canto, donde receptores mediadores del LPT, podría inducir varias estimulaciones sinápticas al IMAN, antes de culminar la fase sensitiva de aprendizaje, sin embargo observo una depresión de estas sinapsis cuando esta fase sensitiva normalmente culmina.
Otra gran pregunta de interés seria ¿Si los cambios neuronales son funcionalmente asociados a cambios con memorización del canto versus el primer ensayo del canto?
En estudios con (Melospiza georgiana) se observo que el número de neuronas del área “X” y del HVC aumentaron en la etapa de memorización, señalando una disminución en la fase sensitivo-motora. Este paradigma podría ser usado para explorar la estrecha relación entre los cambios anatómicos y fisiológicos en las diferentes fases del aprendizaje del canto.
Yaki-Sugiyama y Money (2001), compararon el aprendizaje de la Zebra finches entre los 0 y 30 días post-nacimiento con un canto y entre los 60 y 90 días post-nacimiento con un segundo tutor. Ellos encontraron que aparentemente las neuronas del IMAN alteran su respuesta selectiva bajo estas condiciones. El IMAN recibe una constante estimulación auditiva indirectamente vía el HVC, donde las neuronas del HVC, podrían retener esta selectividad para el mismo canto a través de toda su vida. Mientras que un nuevo reclutamiento de neuronas podrían convertirse en un nuevo tipo de canto. ¿Que ocurriría en especies de amplio repertorio?
Otros estudios señalan que un aislamiento del canto específico de la especie durante un periodo temprano de la fase sensitiva, resulta en una producción de sonido anormal. Aunque el individuo presenta un patrón innato, necesita en una fase temprana sensitiva, la exposición a un tutor específico o una interacción directa entre individuos de la misma especie para convertir el canto innato en uno más complejo en estructura. (Brenowitz y Beecher, 2005). Sin embargo, Leinert y colaboradores (2002) observaron en Acrocephalus sp que el mecanismo de control de la canción en el cerebro, se puede convertir independientemente de la exposición a un tutor.
Cuando se estudia el aprendizaje de este tipo de aves (Serinus canarius) seleccionadas por años para el canto, se aprecia la gran capacidad de adaptar sus neuronas del canto para diferentes estímulos de audición. Son aves que se denominan mentes abiertas (open brain), es decir, tienen la capacidad cada año de incorporar nuevas neuronas al núcleo del canto HVC por “Neurogénesis”. Cuando uno estimula con espectros de cantos o giros ya sean de la misma especie o de otra, se incita la formación y estimulación del núcleo central del canto HVC. Primero se estimulan la formación de neuronas en cantidad y luego en especificidad, es decir, existen neuronas específicas de cada especie que son las primeras en aparecer y que responden al canto propio del individuo, otras neuronas heteroespecificas, se han logrado observar su crecimiento y asociación con giros no propios del individuo, evidentemente estas conexiones se encuentran en menor proporción. Por lo tanto al estimular con cantos o giros se estimula la neurogénesis y las conexiones sinápticas de neuronas específica de la especie y de alguna otras heteroespecificas. Hoy en día todavía esta en estudio lo que se denomina fase sensitiva y motora auditiva del individuo, que particularmente esta ultima es la más importante. Cuando el individuo escucha lo aprendido en la fase sensitiva, interpretando y regulando sus giros (regulación somato sensorial) para generar su propio canto.
Existen evidencias que señalan que cuando ocurre una sobre saturación de la estimulación en la fase sensitiva - motor, con más de tres repertorios heteroespecíficos, no especifico de la especie, se han encontrado una disminución de las conexiones neurológicas para el canto y de la neurogenesis, lo que se traduce en un canto de repertorio mas corto y no bien estructurado.
Neuronas Selectivas del Canto del Individuo, Repertorio de Canto y Plasticidad
Las neuronas en ambas vías motoras y en el AFT son responsables de la estimulación acústica. Algunas neuronas responden en ciertas circunstancias selectivamente al canto del propio individuo. La funcionalidad significativa de estas neuronas selectivas no se tiene todavía muy claro. Estas podría prever cualquier error en la señal, promoviendo cambios en la producción del sonido al detectar la falla entre la retroalimentación de la audición del canto del mismo individuo y el canto memorizado. Podría tener un rol de gran importancia en el aprendizaje del canto y el mantenimiento del mismo. Una segunda función podría ser la percepción específica del canto. Esta función no es necesaria mente específica de estas neuronas, ya que existen un indeterminado número de neuronas que podrían almacenar esta información. En especies con amplios repertorios de cantos, las neuronas individuales responden a tipos de cantos simples o múltiples.
Mooney y colaboradores (2001) encontraron que la mayoría de proyecciones simples de las neuronas del RA al HVC descargan selectivamente por una retroalimentación de un tipo de canto simple, sin embargo las ínter neuronas del HVC responden a todos tipos de sonidos de un canto estereotipado de la especie, pero no a sonidos heteroespecificas.
En especies con amplio repertorio (Serinus canarius), existe el interés de preguntarse, ¿Si las neuronas son específicas no para un canto del ave en particular, sino para un específico tipo de sonido?, producido por el ave o por otras. Seria interesante determinar si el patrón de especificidad encontrado por estos autores en aves de repertorio reducido (Swamp sparrows), ocurren en especies de largo repertorio, tal como el Serinus canarius.
Todavía no se tiene claro sobre el patrón de selectividad neuronal que ocurre en especies de amplio repertorio. Se ha observado en estas especies que las neuronas de un macho responden a la estimulación de otro macho de la misma especie, lo cual explica el comportamiento de varias especies, los cuales usan el canto como medio de interacción y agresividad. Siendo un efecto positivo en el desarrollo de la respuesta neuronal auditiva.
Plasticidad, Diversidad y la Evolución del Aprendizaje Del Canto Adulto
En aves (zebras finches), el aprendizaje del ave es restringido al primer año (limite de edad), conociéndose como aves de aprendizaje cerrado. El Serinus canarius presenta un proceso de aprendizaje similar durante el primer año, siendo capaces de desarrollar nuevos cantos en subsecuentes años como un macho adulto maduro. Estas aves son descritos como aves de aprendizaje abierto, realmente es dificultoso determinar en estas aves sí el nuevo canto desarrollado como adulto envuelve memorización de un nuevo canto modelo o un modelo previamente memorizado.
La carencia de aprendizaje de canto del adulto, ha sido asumida como resultado de una deficiencia de plasticidad del sistema de control de canto de aves de aprendizaje cerrado. Todavía no se tiene claro, ya que el circuito de canto adulto esta caracterizado por una plasticidad extensiva tanto en aves de aprendizaje cerrado como en las especie de aprendizaje abierto. Existen evidencias de algunas especies que pueden manejar estas dos estrategias de aprendizaje. Esto nos hace sugerir que ambas estrategias de aprendizaje pueden ser observadas con diferencias cuantitativas en el grado de plasticidad del canto adulto, más que diferencias cualitativas en la presencia o ausencia de plasticidad.
Cambios Anuales en el Sistema de Canto del Serinus canarius
- Volúmenes del HVC, RA, Área X, y Nxllts.
- Numero de neuronas al HVC.
- Incorporación de nuevas neuronas dentro del HVC.
- Tamaño del cuerpo neuronal en HVC, RA, Área X y IMAN.
- Densidad neuronal en RA y Área X.
- Proyecciones sinápticas y de dendritas al RA.
- Capacidad metabólica de las neuronas del HVC, RA, y Área X.
- Actividad neuro-física de las neuronas del RA.
Existe una diversidad entre las aves cantoras con respecto al circuito de control de canto neural. Esta diversidad ocurre a lo largo de varias dimensiones en el aprendizaje del canto:
1. Tiempo de aprendizaje del canto, conocido como periodo sensitivo de aprendizaje.
2. Numero de cantos aprendidos (tamaño del repertorio)
3. Capacidad del ave en imitar un canto especifico o improvisar por modificación de elementos de sonido, creado noveles cantores.
4. Capacidad de copiar un material tutor único, estrechamente relacionado con los parámetros específicos de la especie o la capacidad especial de copiar cualquier cosa que ellos escuchen.
5. Necesidad de requerir a una exposición temprana de un canto especifico de la especie o el poder desarrollar un canto típico de la especie, cuando son criados en ambientes aislados.
Una explicación de la diversidad, es la dificultad de identificar un único programa de aprendizaje de canto común para todas las aves cantoras. Otro punto de interés es la necesidad general de comparar la retroalimentación auditiva generada por el mismo canto del individuo como un modelo interno.
Existe una estrecha relación entre la neuro-génesis y la plasticidad del canto. Las neuronas son reclutadas por el HVC del ave juvenil (Serinus canarius y Taeniopygia guttata) en una alta tasa, cuando ellos están activamente aprendiendo el canto, para luego producir un canto estereotipado o cristalizado.
Los canarios tienen la capacidad de desarrollar nuevas silabas de sonidos cuando son adultos, incorporando nuevas neuronas dentro del HVC adulto, aumentado en el verano cuando el canto es variable y la adición de silabas es grande. Esta observación nos hace pensar que la adición de neuronas al HVC, esta funcionalmente relacionado con el aprendizaje del canto. Sin embargo aunque la neuro-génesis podría ser necesaria para el aprendizaje del canto, no se tiene todavía claro la relación entre estos dos procesos.
Sí uno intenta correlacionar los cambios en el circuito de canto que ocurren anualmente, con la capacidad somato sensorial de las distintas especies de aves con aprendizaje abierto o cerrado, siempre existen una seria de preguntas que salen a relucir y que se trasforman en futuros objetivos de investigación, demostrando que es un tema y área que necesitan gran interés y dedicación en futuras líneas de investigación.
¿Los cambios anuales de restablecimiento neuronal del HVC son comunes en todas las especies de aves de canto, estando consistentemente asociadas con cambios anuales en el comportamiento de la plasticidad del canto?
¿Los patrones juveniles de la plasticidad neuronal son retenidos o restablecidos anualmente en especies que memorizan o reensayan nuevo canto cuando adulto?
¿En especies con largo repertorio, están las neuronas selectivamente en respuesta única al canto propio del individuo, para un tipo de canto simple o múltiple?
¿Que cambios neuronales ocurren sobre la evolución de las aves de aprendizaje cerrado o abierto en el patrón de aprendizaje de canto?
¿Canto estereotipado, duración y tasa de producción?
¿Será que un mismo tipo de canto para otra ave diferente al sujeto que la produce tan efectivo como la versión propia del individuo, para la estimulación de las neuronas propias del sujeto?
¿En aves con aprendizaje abierto, las neuronas individuales modifican su selectividad para producir cambios en la estructura del canto o nuevamente restablecen el servicio de las neuronas para esta función, es decir para la selectividad?
Herencia ligada al sexo de la audición y canto en el canario Belga Waterslager
El Waterslager tiene menos sensibilidad auditiva a las altas frecuencias, produciendo un canto con más energía a baja frecuencia que el canario de canto silvestre. Un retrocruce en el pedigree entre el Waterslager y las líneas domesticas del canto de tipo silvestre, revelan un patrón consistente de herencia con un factor de mayor efecto localizado en el cromosoma sexual “Z”, afectando ambos cromosomas. Provocando una pobre audición a las altas frecuencias por encima de los 4 Khz y cambios relativos en la energía del espectro de aprendizaje del canto del macho. El umbral de la audición a las 4 Khz fue un significante predictor de la relativa cantidad de energía del canto a los 4 Khz para machos individuales. Una hipótesis para el mecanismo básico de esta correlación entre la audición y las anormalidades del canto esta en la reducción en la habilidad de escuchar sonidos de altas frecuencias, predisponiendo a los machos hacia el aprendizaje de sonidos de bajas frecuencias.
El umbral de la audición en las aves esta típicamente correlacionado con los espectros de frecuencias de su canto. El origen de esta correlación no esta todavía bien entendido, particularmente en especies que aprenden su canto. El canario domestica ha sido seleccionado artificialmente por especificas líneas de cantos, ofreciendo un sistema ideal para la investigación de esta correlación. (Dooling et al, 1971; Guttinger, 1985)
El waterslager exhibe un cambio en el pico de energía de su espectro de aprendizaje del canto a bajas frecuencias, difiriendo marcadamente de lo encontrado en otras razas de cantos del Serinus canarius. Esta línea solo exhibe una pobre capacidad de audición a altas frecuencias relativa a otros canarios, mientras que el umbral de la audición esta debajo de de 2 Khz. normalmente. Aquellos que están por encima de los 2 Khz con un rango de 20 – 40 db de altura se acercan mas al canario de tipo silvestre. (Okanoya y Dooling, 1987)
Esta perdida de la audición a altas frecuencias aparece en los pichones tan temprano como a los 20 días post-n nacimientos, dirigiendo la investigación hacia la perdida y daño de las células de la audición en la papila bacilar del oído interno. (Gleich et al, 1997) El comportamiento consecuente a esta anormalidad anatómica esta en el cambio de rango de menor audición entre 2 a 4 khz en los canarios de tipo silvestres a 1 - 2 Khz en el canario Waterslager (BWS).
Cambios específicos de la línea tanto en el canto como en la audición en el canario podrían estar ligados al sexo (Okanoya et al, 1990; Mundinger, 1995; Mundinger, 1999). En las aves la determinación del sexo esta presente en un revés heterogametico (a diferencia de los mamíferos), en la cual los machos son “ZZ” y las hembras “ZW”. Recientes estudios ha sugeridos que los genes que controlan sexualidad, que selecciona características en este caso de canto, comúnmente ocurren en el cromosoma “Z” (Iyengar et al, 2002; Prince, 2002), a pesar que realidad en las aves este solo comprende el 7 al 10 % del total del genoma. (Bloom et al, 1993).
Efecto del cromosoma “Z” en la sensibilidad auditiva y en la frecuencia de canto de machos con un Retrocruce entre BWS X canario de canto clásico.
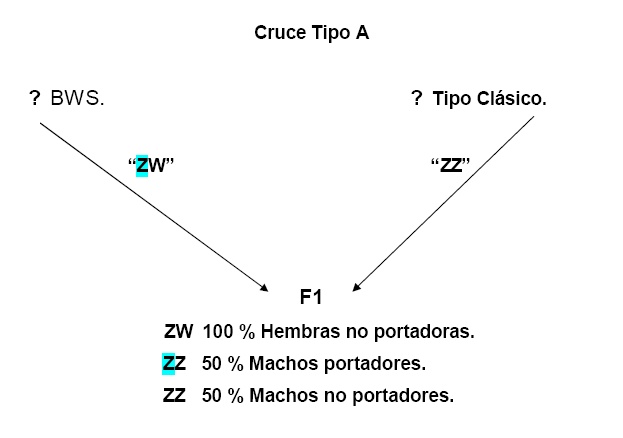
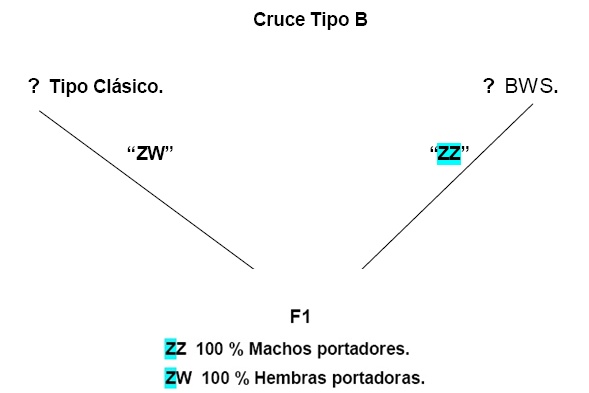
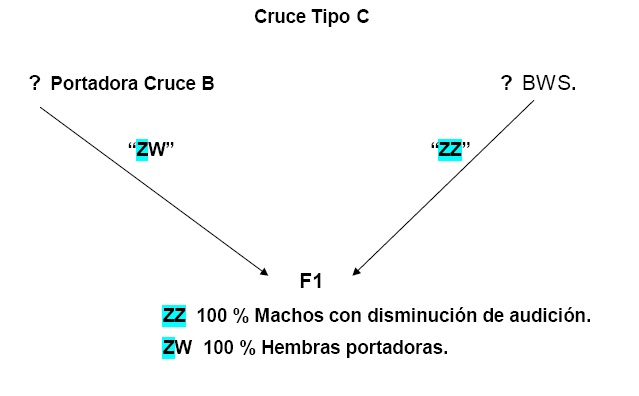
Herencia de la audición y espectro de canto
La presencia de Machos “ZZ”, muestra una marcada disminución del umbral de la audición, observándose un factor genético de mayor efecto localizado en el cromosoma “Z”, con un complemento autosomal indiferente. Con un aparente factor de asociación entre genes sexuales y autosomicos con actividad recesiva. El cromosoma “Z” es estimado por ser el responsable para 87% a 91 % de las diferencias entre 2 clases de audición a los 4 Khz, dependiendo del grado de expresión recesiva asumida. El real significado de la diferencia entre los dos modelos indica que el factor Z-enlace es altamente recesivo.
El patrón de herencia del espectro de energía del canto fue solo consistente con un factor de Z-enlace recesivo. Esto explica entre el 79 al 86 % de la variación del espectro de sonido.
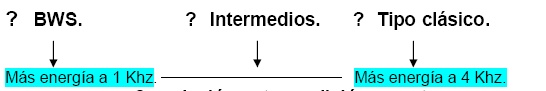
Correlación entre audición y canto
El umbral de audición a 4 khz para individuos machos fue un predictor significativo del espectro de frecuencias de su canto. Esto explica el 49% de la variación de energía de su canto. Una potencial explicación para esta asociación es que esta surja durante el aprendizaje del canto, conduciendo a una percepción pobre de elementos de altas frecuencias del canto de tipo clásico.
Esta parcialidad de percepción podría afectar la fase de memorización (sensitiva), cuando los canarios jóvenes adquieren su capacidad auditiva y la fase motora, cuando ellos atienden competencia de su propio canto con el de otras aves de su misma especie. Esta parcialidad no necesita ser absoluta, por ejemplo, algún aprendizaje de elementos del canto menos perceptibles podrían haber ocurrido bajo condiciones controladas de un proceso de aprendizaje con un tutor. Otra posibilidad, aunque la menos que nos gusta, es observar la asociación entre la audición y el canto que surja entre una correlación genética a través de un gen Pleiotropy (un gen que afecta múltiples rasgos) o una conexión física diferentes genes. Por ejemplo genes que afecten la cadencia de emisión de giros, con mayor apoyo en la discontinuidad.
Conexión Sensorial
La aparente conexión de la audición con los rasgos del canto a través de la percepción sensorial en canarios “BWS” puede tener una general implicación de la evolución de los rasgos de la selección sexual. El aprendizaje del canto de los machos en la aves de canto Oscinas es importante en la preferencia del apareamiento (Searcy y Yasukawa, 1986). En varias especies las hembras aprenden su preferencia sexual por el canto del macho, a través de tempranas exposiciones (Nagle y Kreutzer, 1997; Irwin y Price, 1999). Por lo tanto cambios en la percepción auditiva que afecta el aprendizaje del canto podría potencialmente alterar la expresión del mismo en los machos y la preferencia en las hembras simultáneamente.
Tal conexión sensorial podría proveer un mecanismo para una rápida evolución de un rasgo y un aislamiento reproductivo que nos conllevaría a una selección natural. En donde el aprendizaje del macho y la preferencia de la hembra estarían interconectados en una vía común, dependiente de una salida sensorial mejor que a través de una conexión física de Locis genéticos separados.
Viendo la importancia de este punto, Investigaciones futuras deberían enfocarse en las variaciones que se suceden cuando el macho canta, así como las preferencias de las hembras y la sensibilidad auditiva de un grupo de canarios silvestres, pudiéndonos servir como una prueba hipotética de la conexión sensorial.
El Profesor Clive Catchpole (2003), ha observado como la selección sexual ha formado la evolución de canciones complejas en aves y en última instancia como esta se refleja en la evolución del cerebro mismo. Otros investigadores como Dr. Stefan Leitner, ha estudiado el mecanismo de la opción femenina en pájaros cantores, en donde las hembras seleccionan a machos con canciones mas complejas (repertorios mas grandes), evidenciando que los machos con repertorios amplios, presentan un núcleo mas grande del HVC en las vías del control de la canción. Estos estudios nos permiten inferir en las siguientes preguntas. ¿Será que las hembras seleccionan a los machos con un HVC más grande? O ¿Será que las hembras que más seleccionan son la que tienen los nervios más grandes? En donde la complejidad masculina de la canción se ha desarrollado explotando una respuesta sensorial preexistente en el camino de los nervios femeninos. Leitner y Catchpole (2002), responden a estas preguntas, en donde demostraron que las canarias femeninas reconocen y discrimina mas las canciones del macho de diversa calidad, corroborando la existencia de un núcleo de control de la canción (HVC) mas grande en el cerebro.
Otros estudios por Bernd leisler, utilizando un programa de edición de sonidos, ambiente controlado y exposición de machos en forma aleatoria, se han enfocado en el comportamiento reproductivo de las hembras del serinus canarius, con respecto al repertorio del canto. En donde cabe preguntarse: ¿Que estructura de la canción las hembras encuentran mas atractivas? y ¿Que silabas de determinado giro son las más atractivas?
Spencer y colaboradores estudian las correlaciones existentes entre la calidad y tamaño del repertorio de aves cantoras, con la calidad de la nutrición, carga de parásitos y elevados niveles de corticoides (stress). También hace énfasis en observar, como influye la exposición de las hembras a canciones mas atractivas, ¿Será que producirán huevos mas grandes, con aumento de las testosterona en la yema del huevo?, aumentando el nº de hijos, así como la inclinación hacia un sexo determinado.