Ajustes respiratorios del serinus canarius durante el canto
Publicado el
por Gabriela.
AUTOR: Gilberto Haddad
 La emisión del canto exige notables ajustes en la función respiratoria, la cual debe satisfacer las necesidades pulmonares y al mismo tiempo, proveer adecuadas cantidades y cadencias de flujo de aire en la siringe que permitan la fonación. El cuerpo de las aves no está dividido por un diafragma muscular como ocurre en los mamíferos. En su lugar aparece un delicado tejido situado entre el hígado y el saco aéreo abdominal que tiene muy pocas fibras musculares y no parece jugar un papel relevante en la respiración. Los músculos inspiratorios expanden los sacos aéreos mediante el movimiento ventral del esternón (Fig. 11). Este movimiento es opuesto por los músculos expiratorios, que comprimen los sacos aéreos. En ausencia de canto, la presión expiratoria máxima en el canario es solo de 0,5 a 3,0 cm H2O, pero se incrementa notablemente durante el canto, alcanzando con frecuencia 10 a 15 cm H2O en el saco torácico anterior con máximos medidos de 30 cm para algunas sílabas. Casi la totalidad del canto se produce durante expiraciones. Dos excepciones conocidas son ocasionales sílabas inspiratorias en el canto de algunos pinzones cebra (Goller y Dalley, 2001) y un sonido inspiratorio “woh” asociado a reclamos de palomas. (Gaunt, Gaunt y Casey, 1982)
La emisión del canto exige notables ajustes en la función respiratoria, la cual debe satisfacer las necesidades pulmonares y al mismo tiempo, proveer adecuadas cantidades y cadencias de flujo de aire en la siringe que permitan la fonación. El cuerpo de las aves no está dividido por un diafragma muscular como ocurre en los mamíferos. En su lugar aparece un delicado tejido situado entre el hígado y el saco aéreo abdominal que tiene muy pocas fibras musculares y no parece jugar un papel relevante en la respiración. Los músculos inspiratorios expanden los sacos aéreos mediante el movimiento ventral del esternón (Fig. 11). Este movimiento es opuesto por los músculos expiratorios, que comprimen los sacos aéreos. En ausencia de canto, la presión expiratoria máxima en el canario es solo de 0,5 a 3,0 cm H2O, pero se incrementa notablemente durante el canto, alcanzando con frecuencia 10 a 15 cm H2O en el saco torácico anterior con máximos medidos de 30 cm para algunas sílabas. Casi la totalidad del canto se produce durante expiraciones. Dos excepciones conocidas son ocasionales sílabas inspiratorias en el canto de algunos pinzones cebra (Goller y Dalley, 2001) y un sonido inspiratorio “woh” asociado a reclamos de palomas. (Gaunt, Gaunt y Casey, 1982)
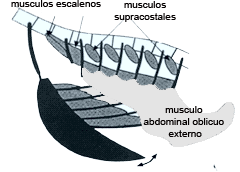
Fig.11 La actividad de los músculos inspiratorios y expiratorios se relaciona con la producción de canto. (a) Alguno de los músculos inspiratorios y uno de los cuatro músculos expiratorios abdominales. (Tomado de Suthers y Zollinger, 2004)
Durante el canto el modelo respiratorio motor debe ser coordinado con el de la siringe. Potencialmente esto puede implicar hasta a 11 músculos respiratorios principales que participan en la respiración normal además de 8 músculos adicionales que registran actividad en estas respiraciones especiales. (Fedde, 1987) Hasta la fecha solo han sido registrados electromiogramas durante el canto de unos pocos músculos expiratorios abdominales (fundamentalmente del músculo oblicuo abdominal externo y del transverso) y de dos músculos inspiratorios torácicos, el escaleno y lod musculos elevadores costales (Fig.11). El papel del resto de los músculos respiratorios durante el canto se desconoce. En pinzones cebra, cardenales y cow birds la amplitud del EMG durante el canto se incrementa entre 5 y 12 partes comparada con respiración normal, probablemente manifestando mayor actividad muscular y participación de más músculos (Wild, Goller y Suthers, 1998; Suthers, Goller y Wild, 2002). La actividad eléctrica de los músculos expiratorios e inspiratorios no se superpone en el tiempo. (Wild, Goller y Suthers, 1998) La actividad de los músculos expiratorios no aparece lateralizada. La similar amplitud de los EMG en ambos lados de la siringe, con independencia del lado que está produciendo sonido, sugiere control motor bilateral. Los músculos expiratorios modulan su actividad en ambos lados de la misma manera al generar modelos silábicos específicos de presión y flujo de aire, incluso cuando la fonación es unilateral. (Goller y Suthers, 1999)
El modelo temporal respiratorio cambia marcadamente durante el canto, en función de la cadencia silábica del canto, las aves canoras emplean uno de los dos siguientes modelos respiratorios. A cadencias silábicas moderadas toman una breve mini-inspiración tras cada sílaba emitida para reemplazar el volumen de aire exhalado en la producción del sonido (Hartley y Suthers 1989; Vicario, 1991). La duración de las sílabas generalmente tiene una relación inversa con la cadencia de repetición. Puesto que las sílabas largas emplean más aire, las mini-respiraciones han de ser también más prolongadas. En canarios Malinois, por ejemplo, una sílaba de 119ms. de duración cantada a un ritmo de 6,5 sílabas/segundo necesitó 0,25ml de aire para ser producida, lo cual fue también el volumen de su mini-inspiración asociada. El volumen correspondiente a una sílaba de 11ms. cantada con una cadencia de 30 sílabas/segundo fue de unos 0,04ml. (Hartley y Suthers, 1989)
Con el empleo de mini-respiraciones un ave podría, en teoría, mantener un volumen respiratorio casi constante durante el canto. Esto puede proporcionar importantes ventajas en la producción de precisos y estereotipados movimientos respiratorios necesarios durante la vocalización, puesto que los músculos respiratorios podrían operar lo más cerca posible de su estado de descanso, pudiendo, prever la fuerza elástica de retroceso del tórax. La dirección del retroceso elástico del esternón es ambivalente: cuando el volumen de aire en los sacos aéreos es pequeño, el retroceso elástico de tórax suplementa el esfuerzo inspiratorio de los músculos al devolver al esternón a su posición neutra de descanso; sin embrago si el volumen en los sacos es grande, la fuerza de retroceso actúa en sentido contrario y suplementa el esfuerzo expiratorio; Un volumen respiratorio constante durante el canto permitiría oscilar al esternón cerca de su punto neutro de descanso de forma que el retroceso elástico pudiera suplementar la acción tanto de los músculos inspiratorios como expiratorios, tal vez permitiendo, además, más rápidas mini-respiraciones y cadencias de repetición silábica más elevadas. (Suthers y Zollinger, 2004)
Es también posible que la sintaxis del canto esté parcialmente influida por las necesidades respiratorias. A altas cadencias silábicas el volumen de las mini-respiraciones es más pequeño que la capacidad de la tráquea. (Hartley y Suthers 1989) No sabemos si estas mini-respiraciones proporcionan oxígeno a los pulmones. Es posible que el ave ajuste la sintaxis de su canto a sus necesidades respiratorias sin verse en la necesidad de interrumpir el canto para una respiración prolongada, introduciendo periódicamente fraseos de baja cadencia y sílabas prolongadas que permitan largas mini-respiraciones mediante las que se puedan ventilar los pulmones. (Suthers y Zollinger, 2004)
Para fraseos interpretados a muy altas cadencias silábicas las aves emplean un patrón respiratorio motor diferente denominado “expiración pulsátil”. La cadencia silábica se incrementa al eliminarse las mini-respiraciones con el costo de tener que limitar la duración del fraseo cuando las reservas de aire u oxígeno queden agotadas. Durante una frase emitida con expiración pulsátil, la presión subsiríngea de los sacos aéreos se mantiene gracias a la actividad de los músculos expiratorios. Generalmente uno de los lados de la siringe permanece cerrado durante toda la frase y el otro lado se abre periódicamente para permitir la salida de una pequeña cantidad de aire que produce la sílaba. (Hartley y Suthers 1989; Hartley, 1990); Excepcionalmente ambos lados de la siringe contribuyen en la emisión de la sílaba. La cadencia de repetición a la cual las aves cambian de mini-respiraciones a respiración pulsátil disminuye con el incremento de la masa corporal, siendo de alrededor de 30 sílabas/segundo en canarios Malinois de 18gr. de peso y de alrededor de 10 sílabas/segundo en mockingbirds de unos 50gr. Esta relación inversa probablemente se deba a que la mayor masa corporal implica una mayor inercia de las paredes torácicas y abdominales que deben oscilar a la frecuencia marcada por la respiración. (Suthers y Zollinger, 2004)
Zollinger y Suthers (2004), comprobaron esta hipótesis educando un ejemplar de northern mockingbird (Mimus polyglottos) con fraseos de canario de alta repetición silábica de 22 sílabas/segundo, comprobando que el ave no era capaz de efectuar mini-respiraciones entre las sílabas. En vez de esto dividió el fraseo en grupos de sílabas. Cada grupo fue interpretado empleando expiración pulsátil efectuando una pausa entre grupo y grupo para una inspiración. A cadencias de repetición inferiores a 11 sílabas/segundo se producen más largos intervalos entre sílaba y sílaba y el mockingbird fue capaz de emplear mini-respiraciones en los fraseos igualando al canario en la duración de éstos. (Suthers y Zollinger, 2004)
El canto del canario que sirvió como tutor es un trino continuo que dura cuatro segundos. A la cadencia emitida, de 22 sílabas/segundo, el canario emplea un patrón respiratorio de mini-respiraciones. El mucho mayor mockingbird no puede interpretar dicha cadencia silábica usando mini-respiraciones, sino que rompe el fraseo en pequeños segmentos cada uno de los cuales se compone de varias sílabas.
Cada fragmento fue interpretado empleando un patrón respiratorio pulsátil. Dentro de cada segmento el mockingbird copió adecuadamente la cadencia de repetición del tutor (22 sílabas/segundo), pero sin mini-respiraciones para reponer el aire utilizado, se vio obligado a interrumpir periódicamente el trino para abrir el lado no fonatorio y realizar una inspiración. La duración total del trino interrumpido es un segundo menor que la del tutor. A menores cadencias de repetición (por debajo de 11 sílabas/segundo), los mockingbirds fueron capaces de usar modelos de mini-respiración, al igual que los canarios, pudieron interpretar más largos fraseos ininterrumpidos (hasta de 9 segundos). (Suthers y Zollinger, 2004)